Defining petal cell identity layer-by-layer
Humberto Herrera-Ubaldo

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
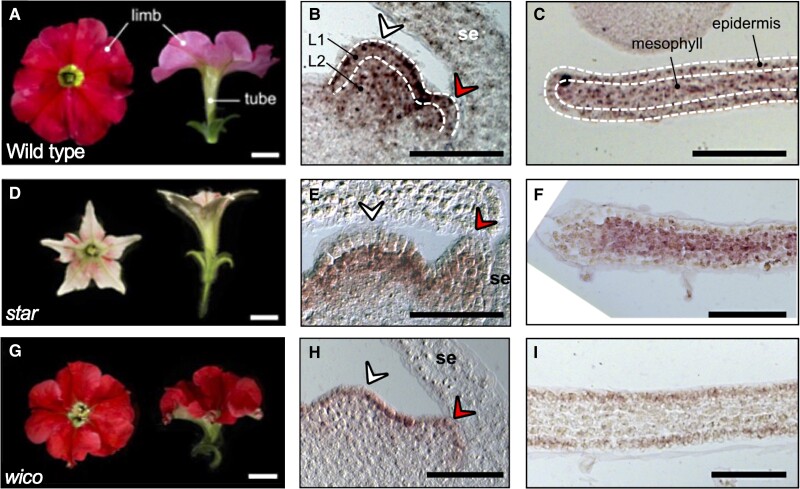 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Molecular Biology Research · Plant Reproductive Biology · Plant Gene Expression Analysis
Flowers' great diversity and beauty are explained by extensive variation in petal color, shape, and pattern. The combination of colors and sizes in the flower organs is key to attraction and interaction with pollinators, hence affecting reproduction. At the molecular level, the formation of floral organs is guided by the combinatorial action of transcription factors as illustrated by the classical ABC model (Coen and Meyerowitz 1991). Petal identity is controlled by the B-class proteins, for example, APETALA3 and PISTILLATA in Arabidopsis or DEFICIENS and GLOBOSA in Antirrhinum (Purugganan et al. 1995); then other transcription factors acting downstream control pigmentation, color patterns, and other special features in the petals (Kellenberger et al. 2023). Our understanding of the molecular mechanisms guiding floral diversity is limited.
In this issue of The Plant Cell, Mathilde Chopy and colleagues (Chopy et al. 2023) dissected the function of the B-class protein DEFICIENS during petal development in petunia flowers and identified cell layer–specific mechanisms that guide the formation of the corolla tube and limb. In Petunia, there are 4 B-class proteins, PhDEFICIENS and PhTM6 (DEF/AP3 clade) and PhGLO1 and PhGLO2 (GLO/PI clade). Double mutants in B-class genes have the classical B-function mutant phenotype: petals are converted into sepals and stamens into carpels. In contrast, the functions of the DEF/AP3 clade have diverged. PhDEF works in the petal, whereas PhTM6 (in combination with C-class proteins) functions in stamen development. The single mutant phdef displays homeotic conversion of the petal whorl into sepals; in contrast, stamen development is unaltered (Rijpkema et al. 2006).
The petunia flower has 5 fused petals with 2 well-defined domains: a flat, wide, and colorful limb and a long tubular structure called the tube (see Fig.); variations in tube length can affect its interaction with pollinators. The phdef-151 allele is a null mutation caused by the insertion of the dTph1 transposon in the PhDEF locus. In the phdef-151 mutant, the petals are converted into sepals; however, in some cases, the authors observed recovery as well as a restoration of petal development in some branches of the phdef-151 plants. They classified the recovery events into 2 classes, wico and star, with contrasting features. The wico mutant displayed nearly normal corolla development, but the tube was underdeveloped, showing a reduction of about 3-fold in length. On the other hand, the star mutant showed strongly reduced lateral expansion, causing a 5-fold decrease in the limb area; the tube displayed a slight reduction in length. In summary, the tube and limb development programs could be uncoupled.
The recovery of petal formation in some presumed phdef-151 mutants suggested that phDEF activity was reactivated, probably mediated by transposon movement out of the PhDEF locus. The rearrangement events were analyzed by sequencing the PhDEF locus in the wico and star mutants; surprisingly, they found similar sequences, either wild type or sequences with +6 base pairs producing 2 extra amino acids in PhDEF. Because there was no difference at the sequence level, the authors analyzed the PhDEF transcript distribution using in situ mRNA hybridization. The PhDEF transcript has a wide distribution across the different cell layers in the wild-type flower meristems (L1–L3). However, in the wico mutant, the signal was detected only in the L1; conversely, in the star mutant, the transcript was detected in the L2 and L3. These results indicate the function of PhDEF can be spatially modular: its transcript presence in the L1 drives the development of the limb, whereas its presence in the L2 drives the development of the tube. In summary, the expression of PhDEF in the petal epidermis is the main driver of limb morphogenesis and controls features such as growth, shape, and pigmentation, whereas mesophyll drives tube morphogenesis.
Next, analyses were conducted to study cell identity across the petal cell layers. The wild-type adaxial limb cells are conical, whereas the tube cells are elongated. In wico, the limb cells remain conical but bigger, whereas cells in the tube are similar in shape but shorter. Cells in the star limb are rounder, and cells in the tube are similar but shorter. Interestingly, in star the epidermal cells in the limb have an intermediate identity between petal and sepals. The layer-specific expression of PhDEF in wico or star lines is a good system to study the function of PhDEF in the cells where it is expressed (cell-autonomous function) and the role in the neighbor layers (non-cell autonomous).
Finally, to look at the processes downstream of PhDEF, the authors conducted gene expression analysis through petal development in the wild-type, phdef-151, wico, and star lines. They focused on differentially expressed genes related to the production of pigments. Comparison between the phdef-151 or star and the wico expressed genes indicated a strong downregulation of genes in the anthocyanin biosynthesis pathway. Interestingly, they identified some genes encoding major regulators of petal pigmentation, in particular, some members of the MBW (MYB, bHLH, WD40) complex. They found the predicted PhDEF DNA-binding sequence in the ANTHOCYANIN1 (AN1) and AN2 regulatory regions. The binding events were tested using electro-mobility shift assays; in vivo analysis found PhDEF directly binds to the terminator region of AN2, indicating PhDEF could directly control pigmentation in the petal limb epidermis.
In summary, the B-class protein PhDEFICIENS has layer-specific functions during petal development. In the epidermis, PhDEF has a role in the control of petal pigmentation, conical cell shape, and limb growth, whereas the expression in the mesophyll directs tube growth.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Coen ES , Meyerowitz EM. The war of the whorls: genetic interactions controlling flower development. Nature. 1991:353(6339):31–37. 10.1038/353031 a 01715520 · doi ↗ · pubmed ↗
- 2Chopy M , Cavallini-Speisser Q, Chambrier P, Morel P, Just J, Hugouvieux V, Rodrigues Bento S, Zubieta C, Vandenbussche M, Monniaux M. Cell layer-specific expression of the homeotic MADS-box transcription factor Ph DEF contributes to modular petal morphogenesis in petunia. Plant Cell. 2024:36(2):324–345. 10.1093/plcell/koad 258PMC 1082731337804091 · doi ↗ · pubmed ↗
- 3Kellenberger RT , Ponraj U, Delahaie B, Fattorini R, Balk J, Lopez-Gomollon S, Müller KH, Ellis AG, Glover BJ. Multiple gene co-options underlie the rapid evolution of sexually deceptive flowers in Gorteria diffusa. Curr Biol.2023:33(8):1502–1512.e 8. 10.1016/j.cub.2023.03.00336963385 · doi ↗ · pubmed ↗
- 4Purugganan MD , Rounsley SD, Schmidt RJ, Yanofsky MF. Molecular evolution of flower development: diversification of the plant MADS-box regulatory gene family. Genetics. 1995:140(1):345–356. 10.1093/genetics/140.1.3457635298 PMC 1206560 · doi ↗ · pubmed ↗
- 5Rijpkema AS , Royaert S, Zethof J, van der Weerden G, Gerats T, Vandenbussche M. Analysis of the Petunia TM 6 MADS-box gene reveals functional divergence within the DEF/AP 3 lineage. Plant Cell. 2006:18(8):1819–1832. 10.1105/tpc.106.04293716844905 PMC 1533978 · doi ↗ · pubmed ↗