Excitable solitons: Annihilation, crossover, and nucleation of pulses in mass-conserving activator-inhibitor media
Arik Yochelis, Carsten Beta, Nir S. Gov

TL;DR
This paper reveals that in mass-conserving activator-inhibitor systems, excitable pulses can undergo soliton-like crossover and nucleation upon collision, contrasting with traditional annihilation behavior.
Contribution
It demonstrates that mass conservation leads to novel collision outcomes of excitable pulses, supported by a model of intracellular actin waves.
Findings
Colliding pulses can exhibit crossover and nucleation.
Mass conservation enables robust, long-lived wave dynamics.
Results explain the ubiquity of actin waves in cells.
Abstract
Excitable pulses are among the most widespread dynamical patterns that occur in many different systems, ranging from biological cells to chemical reactions and ecological populations. Traditionally, the mutual annihilation of two colliding pulses is regarded as their prototypical signature. Here we show that colliding excitable pulses may exhibit soliton-like crossover and pulse nucleation if the system obeys a mass conservation constraint. In contrast to previous observations in systems without mass conservation, these alternative collision scenarios are robustly observed over a wide range of parameters. We demonstrate our findings using a model of intracellular actin waves since, on time scales of wave propagations over the cell scale, cells obey the conservation of actin monomers. The results provide a key concept to understand the ubiquitous occurrence of actin waves in cells,…
Click any figure to enlarge with its caption.
 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3 Figure 4
Figure 4 Figure 5
Figure 5 Figure 6
Figure 6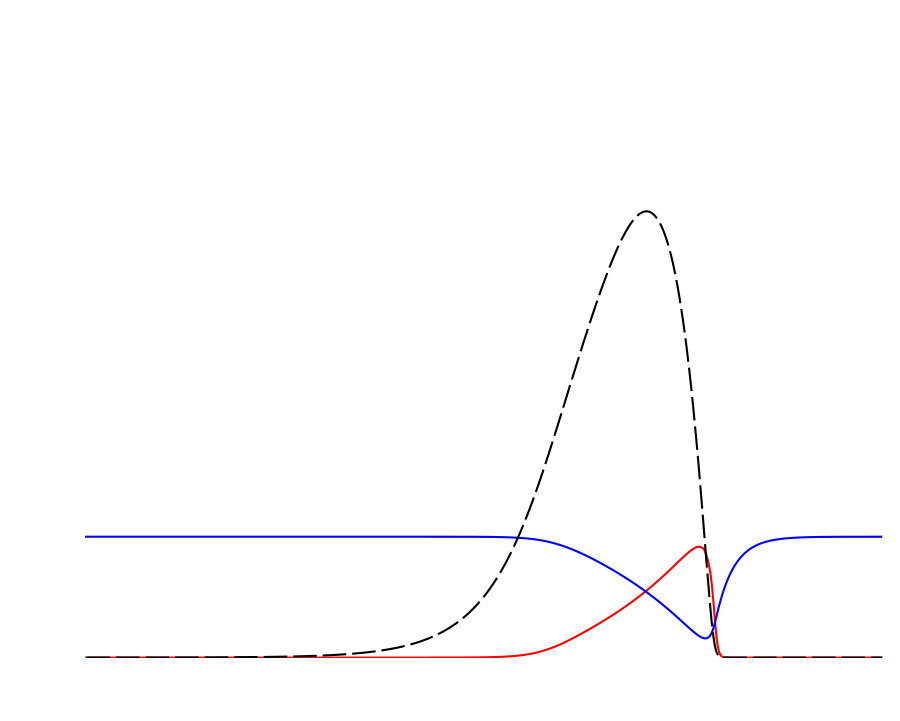 Figure 7
Figure 7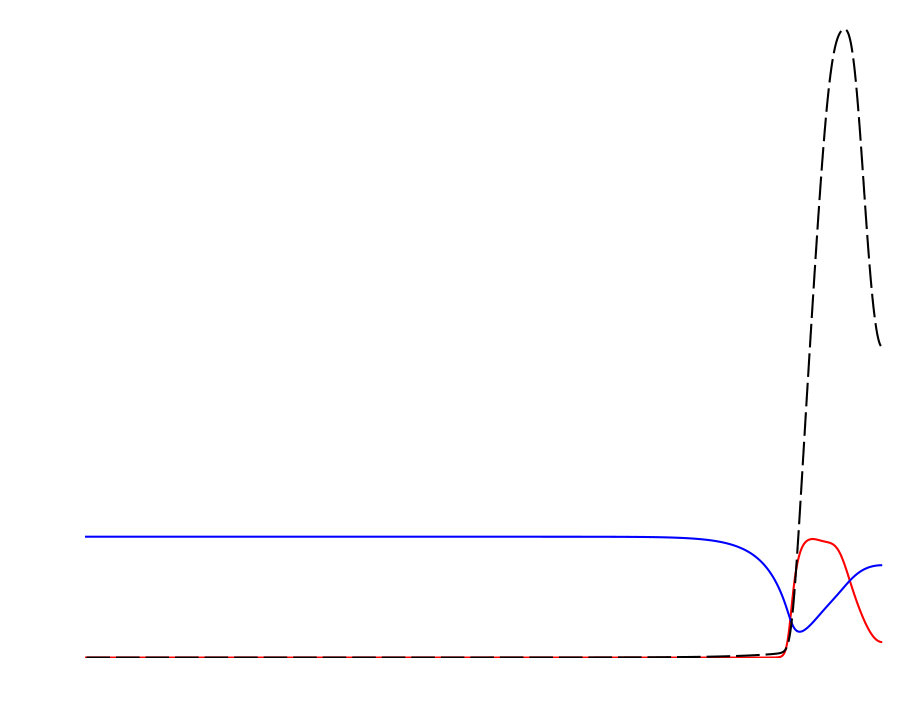 Figure 8
Figure 8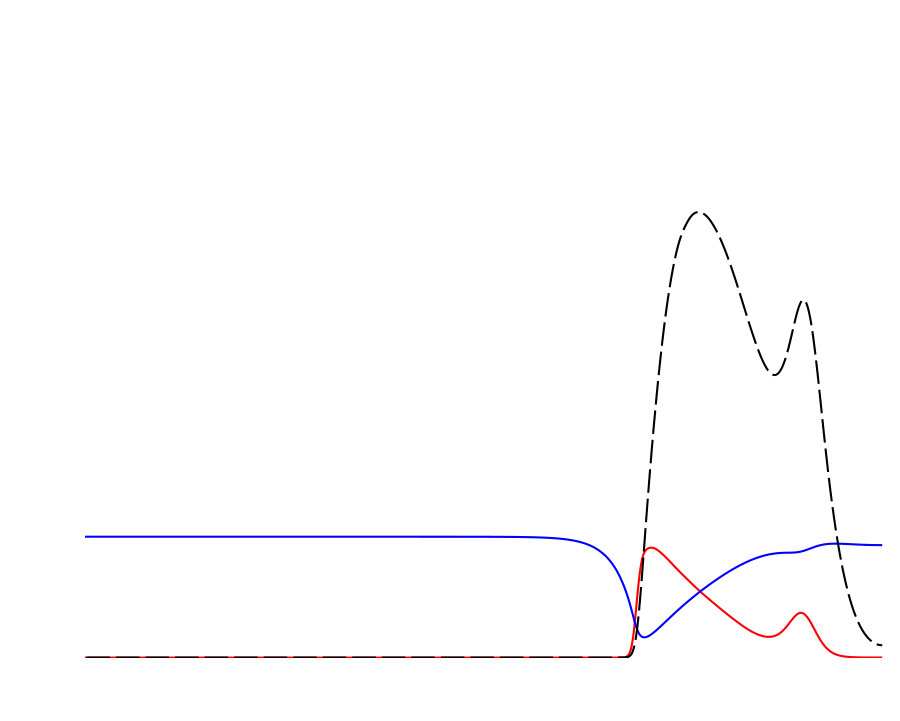 Figure 9
Figure 9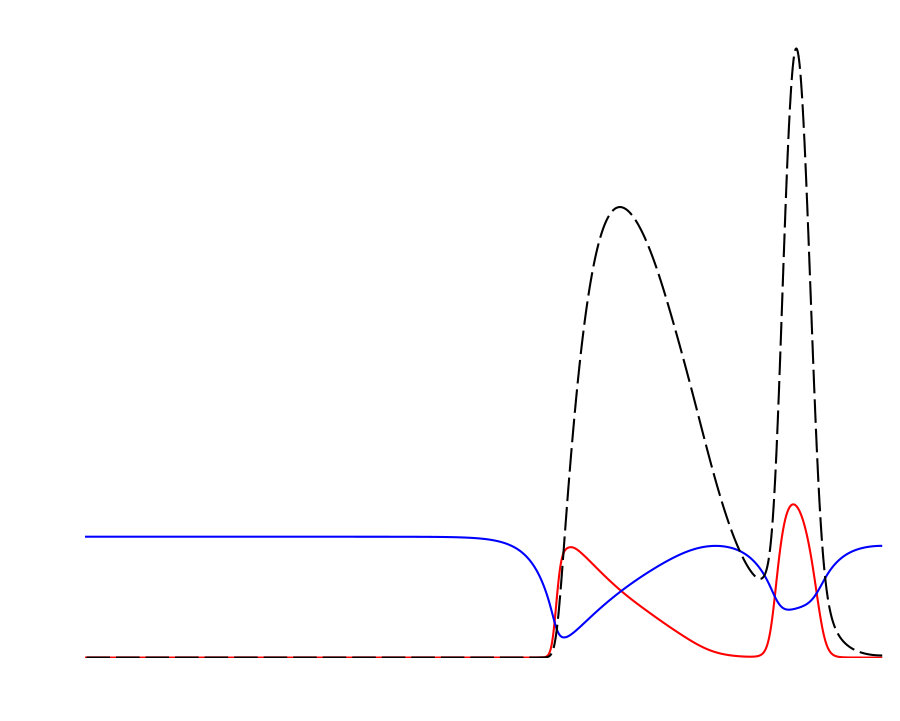 Figure 10
Figure 10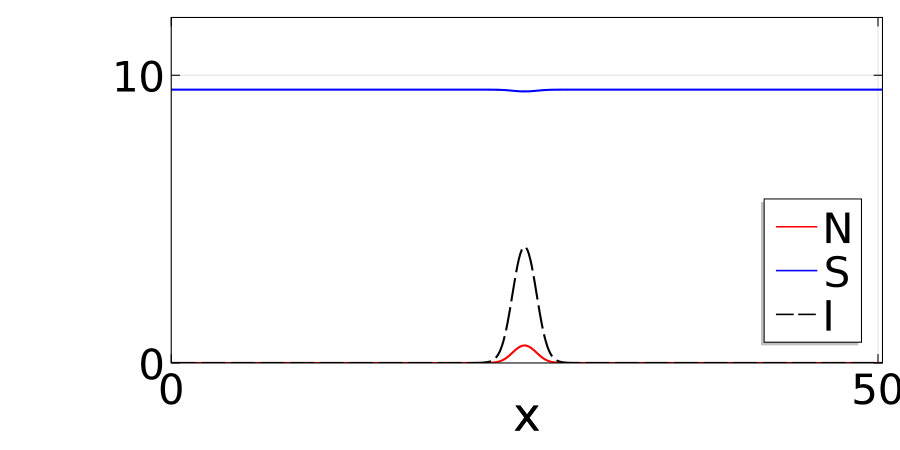 Figure 11
Figure 11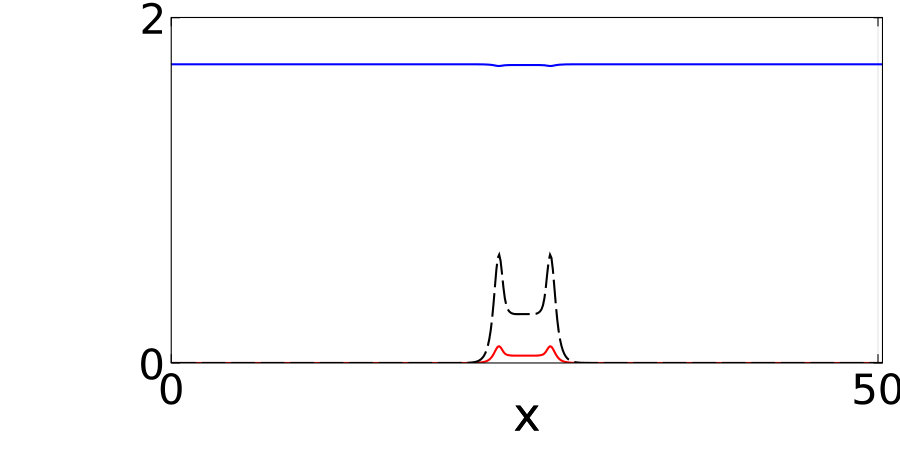 Figure 12
Figure 12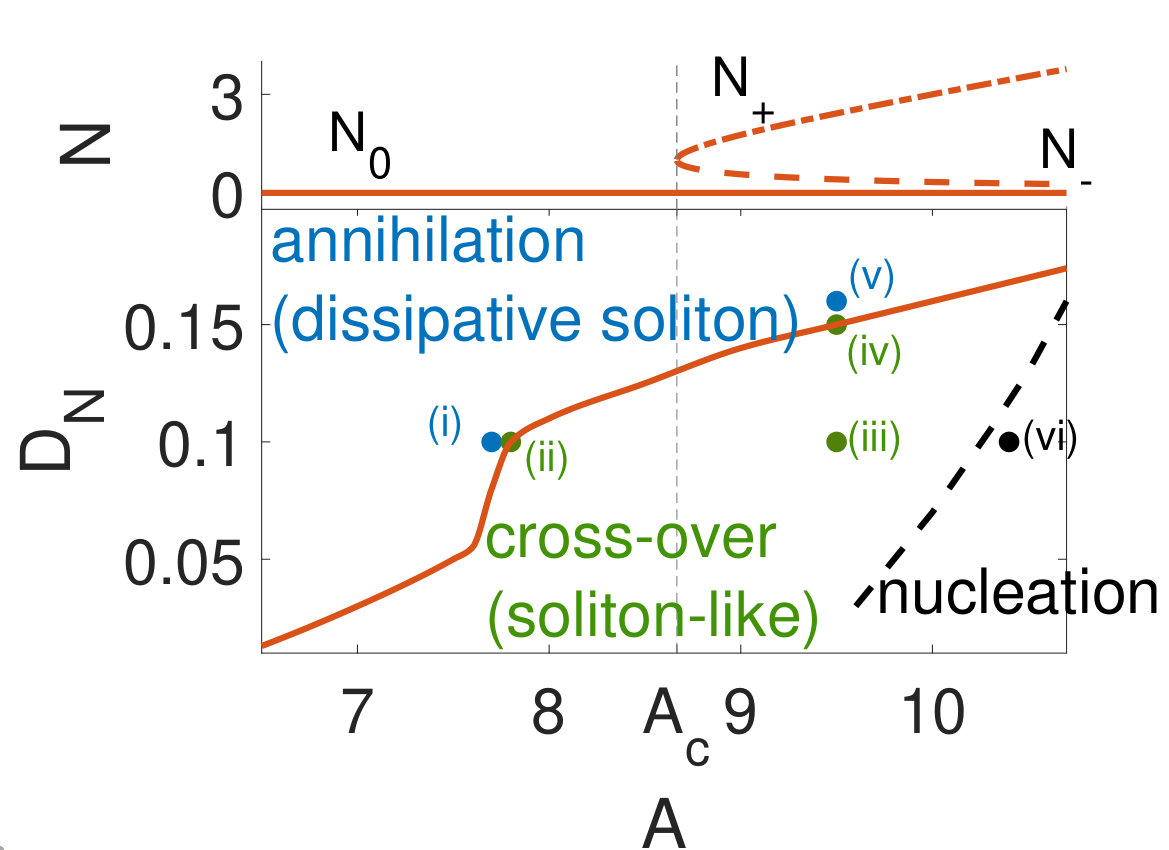 Figure 13
Figure 13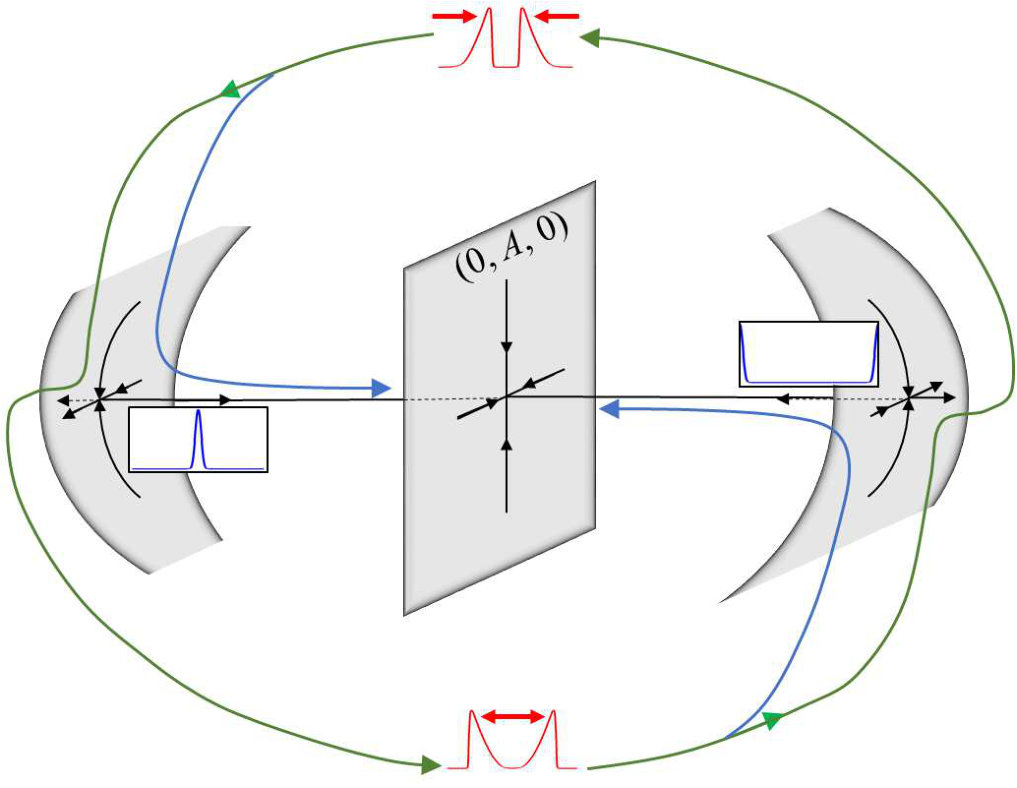 Figure 14
Figure 14Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Excitable solitons: Annihilation, crossover, and nucleation of pulses in mass-conserving activator-inhibitor media
Arik Yochelis
Department of Solar Energy and Environmental Physics, Blaustein Institutes for Desert Research (BIDR), Ben-Gurion University of the Negev, Sede Boqer Campus, Midreshet Ben-Gurion 8499000, Israel
Department of Physics, Ben-Gurion University of the Negev, Be’er Sheva 8410501, Israel
Carsten Beta
Institute of Physics and Astronomy, University of Potsdam, 14476 Potsdam, Germany
Nir S. Gov
Department of Chemical and Biological Physics, Weizmann Institute of Science, Rehovot 76100, Israel
Abstract
Excitable pulses are among the most widespread dynamical patterns that occur in many different systems, ranging from biological cells to chemical reactions and ecological populations. Traditionally, the mutual annihilation of two colliding pulses is regarded as their prototypical signature. Here we show that colliding excitable pulses may exhibit soliton-like crossover and pulse nucleation if the system obeys a mass conservation constraint. In contrast to previous observations in systems without mass conservation, these alternative collision scenarios are robustly observed over a wide range of parameters. We demonstrate our findings using a model of intracellular actin waves since, on time scales of wave propagations over the cell scale, cells obey conservation of actin monomers. The results provide a key concept to understand the ubiquitous occurrence of actin waves in cells, suggesting why they are so common, and why their dynamics is robust and long-lived.
I Introduction
The study of propagating solitary pulses is a cross-disciplinary field of research with important applications in biological, chemical, and physical systems Akhmediev and Ankiewicz (2008); Purwins et al. (2010). Solitary waves are commonly distinguished by their collision properties Scott et al. (1973); Scott (1975): solitons if after collision of two pulses, two pulses emerge (particle-like identity) and dissipative solitons or excitable pulses if they are annihilated. While solitons are often discussed in the context of conservative media, excitable pulses typically arise in dissipative systems that contain auto-catalytic or enzymatic terms Meron (1992); Cross and Hohenberg (1993). Owing to their universal properties, they emerge over a wide range of scales, e.g., in surface reactions Rotermund et al. (1991); von Oertzen et al. (1998), gas discharge plasmas Bode and Purwins (1995), intracellular actin dynamics Allard and Mogilner (2013); Deneke and Di Talia (2018), cardiac rhythms Karma (2013), and neuroscience Izhikevich (2007).
Annihilation of excitable pulses after a collision is well understood and recognized as paramount for electrophysiological function, as it would be impossible to maintain directionality and rhythmic behavior under the reflection of action potentials Alonso et al. (2016). However, in several experimental Rotermund et al. (1991); von Oertzen et al. (1998); Santiago-Rosanne et al. (1997); Willebrand et al. (1992); Shrivastava et al. (2018) and theoretical cases Tuckwell (1979); Kosek and Marek (1995); Mornev et al. (1996); Aslanidi and Mornev (1999); Nishiura et al. (2005); Tsyganov et al. (2007); Lautrup et al. (2011); Young et al. (2019), it was shown that also soliton-like behavior can be observed in dissipative reaction–diffusion (RD) media — a finding that is typically restricted to a narrow range in parameter space and, to date, is considered as an exotic exception to the prototypical annihilation of excitable pulses.
Here we show that soliton-like behavior can robustly emerge in excitable RD media if they obey a mass-conservation constraint. In contrast to previous cases without mass conservation, no fine-tuning of parameters is needed to observe crossover and pulse nucleation upon collision. In particular, neither proximity to an oscillatory onset Argentina et al. (2000) nor non-local interactions Krischer and Mikhailov (1994); Mimura and Kawaguchi (1998); Coombes and Owen (2007) nor cross–diffusion Tsyganov et al. (2003) are required. To underline this paradigmatic shift in our understanding of excitable media, we term pulses in this regime as excitable solitons.
Our results are particularly important to understand dynamical patterns in biological cells, where mass conservation is often a dominant feature. While cells are open systems that grow and divide, the total copy number of proteins such as actin monomers, vary over time-scales that are long compared to the time-scale it takes actin waves to propagate over the cell scale. A prominent example are intracellular actin-membrane waves Gerisch et al. (2004); Bretschneider et al. (2009); Gerhardt et al. (2014) that are associated with fundamental cellular functions and appear in many cell types Beta and Kruse (2017); Inagaki and Katsuno (2017), including Dictyostelium cells Vicker et al. (1997); Gerisch et al. (2004); Gerhardt et al. (2014), neutrophils Weiner et al. (2007), and fish keratocytes Barnhart et al. (2011). We, therefore, demonstrate our findings using the generalized version of a recently developed RD model with mass conservation that successfully describes the dynamics of wave-like actin polymerization in circular dorsal ruffles Bernitt et al. (2017). We find rich dynamics of pulses upon collision, exhibiting not only the common regime of annihilation, but also soliton-like crossover and pulse nucleation over a wide range of parameters.
II Mass-conserved RD model
We start with an RD case model that was formulated to study front dynamics of circular dorsal ruffles (CDR) Bernitt et al. (2017), which are waves of actin polymerization that propagate on the dorsal side of the cell membrane. We reduce this model to a simpler version that includes filamentous actin and an inhibitor of actin polymerization. The three species in this minimal version are: (i) Polymerized actin filaments (F-actin) that are organized in a network (dendritic-like) morphology, , (ii) Actin monomers (G-actin) , and (iii) an actin polymerization inhibitor, . In accordance with the CDR model, we employ actin mass-conservation Bernitt et al. (2017):
[TABLE]
where is the spatial domain size and is constant. In comparison with the CDR model, we have excluded from the current model the additional reservoirs of polymerized actin in the cortex and in stress fibers. Adding them complicates the analysis and does not qualitatively change the nature of the solitary pulses, which are the focus of this study.
The continuum model in its dimensionless form reads Bernitt et al. (2017):
[TABLE]
Eqs. 1a,1b describe the auto-catalytic polymerization process, converting monomers to filaments, which is inhibited by the presence of , and with a constant rate of depolymerization. Eq. 1c describes the recruitment of the inhibitor to the filamentous actin. The hierarchy of diffusion coefficients, along the membrane, is such that the monomers diffuse the fastest, while the effective diffusion of polymerized actin is slower and mostly occurs by the polymerization activity. The inhibitor diffuses the slowest as it is adsorbed to the membrane Bernitt et al. (2017): . In fact, is not essential for what follows, but we keep it as it makes the comparison to the FitzHugh-Nagumo (FHN) model Argentina et al. (2000) transparent. In addition, we chose Bernitt et al. (2017), but this is not essential. We employ Neumann (no-flux) boundary conditions (BC), while similar results (not surprisingly) are obtained with periodic BC.
III Linear stability analysis of uniform solutions
Our interest is in pulses, a situation that requires linear stability of a uniform solution. Eqs. 1 admit three uniform solutions :
[TABLE]
[TABLE]
where, and superscript stands for transpose. Beyond the saddle–node (fold) bifurcation at
[TABLE]
see Fig. III(a), top panel, the solutions appear: is unstable by definition, while linear stability analysis of to uniform perturbations shows that it is also unstable to Hopf oscillations, already from the saddle–node bifurcation point.
Next, we check linear stability of to nonuniform perturbations on an infinite domain Cross and Hohenberg (1993),
[TABLE]
where, is the growth rate of perturbations that are characterized by wavenumbers and c.c. stands for complex conjugate. We find that solution continues to be linearly stable and does not lie in a proximity to any linear oscillatory instability since all parameters are positive, with dispersion relations:
[TABLE]
While the solution was found to be unstable to uniform perturbations, we find that it is unstable also to traveling waves, i.e., non-vanishing imaginary part of its eigenvalue . However, these traveling waves are beyond the scope of our interest here and therefore, not shown.
Notably, the signature of mass-conservation is reflected in the persistence of the neutral mode (Eq. 2b), which indicates a respective mass exchange between and . This property is absent in the typical RD system without mass-conservation, e.g., FHN Argentina et al. (2000), and in what follows, we show that it plays an essential role during the collision of two counter propagating pulses, as shown in Fig. III.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Akhmediev and Ankiewicz (2008) N. Akhmediev and A. Ankiewicz, Dissipative Solitons: From Optics to Biology and Medicine , Vol. 751 (Springer Science & Business Media, 2008).
- 2Purwins et al. (2010) H.-G. Purwins, H. Bödeker, and S. Amiranashvili, Advances in Physics 59 , 485 (2010).
- 3Scott et al. (1973) A. C. Scott, F. Chu, and D. W. Mc Laughlin, Proceedings of the IEEE 61 , 1443 (1973).
- 4Scott (1975) A. C. Scott, Rev. Mod. Phys. 47 , 487 (1975).
- 5Meron (1992) E. Meron, Physics Reports 218 , 1 (1992).
- 6Cross and Hohenberg (1993) M. C. Cross and P. C. Hohenberg, Rev. Mod. Phys. 65 , 851 (1993).
- 7Rotermund et al. (1991) H. Rotermund, S. Jakubith, A. von Oertzen, and G. Ertl, Physical Review Letters 66 , 3083 (1991).
- 8von Oertzen et al. (1998) A. von Oertzen, A. S. Mikhailov, H. H. Rotermund, and G. Ertl, The Journal of Physical Chemistry B 102 , 4966 (1998).