Stiffening and Inelastic Fluidization in Vimentin Intermediate Filament Networks
Anders Aufderhorst-Roberts, Gijsje H. Koenderink

TL;DR
This study investigates how vimentin intermediate filament networks respond to large strains, revealing a rate-dependent softening phenomenon called inelastic fluidization caused by transient crosslinking, which enhances understanding of cell mechanics.
Contribution
It demonstrates that inelastic fluidization in vimentin networks is driven by transient crosslinks, providing new insights into their nonlinear rheology and mechanical resilience.
Findings
Vimentin networks exhibit elastic stress-stiffening.
Identification of a characteristic yield stress $\sigma^*$.
Rate-dependent softening (inelastic fluidization) linked to transient crosslinking.
Abstract
Intermediate filaments are cytoskeletal proteins that are key regulators of cell mechanics, a role which is intrinsically tied to their hierarchical structure and their unique ability to accommodate large axial strains. However, how the single-filament response to applied strains translates to networks remains unclear, particularly with regards to the crosslinking role played by the filaments' disordered "tail" domains. Here we test the role of these noncovalent crosslinks in the nonlinear rheology of reconstituted networks of the intermediate filament protein vimentin, probing their stress- and rate- dependent mechanics. Similarly to previous studies we observe elastic stress-stiffening but unlike previous work we identify a characteristic yield stress , above which the networks exhibit rate-dependent softening of the network, referred to as .…
Click any figure to enlarge with its caption.
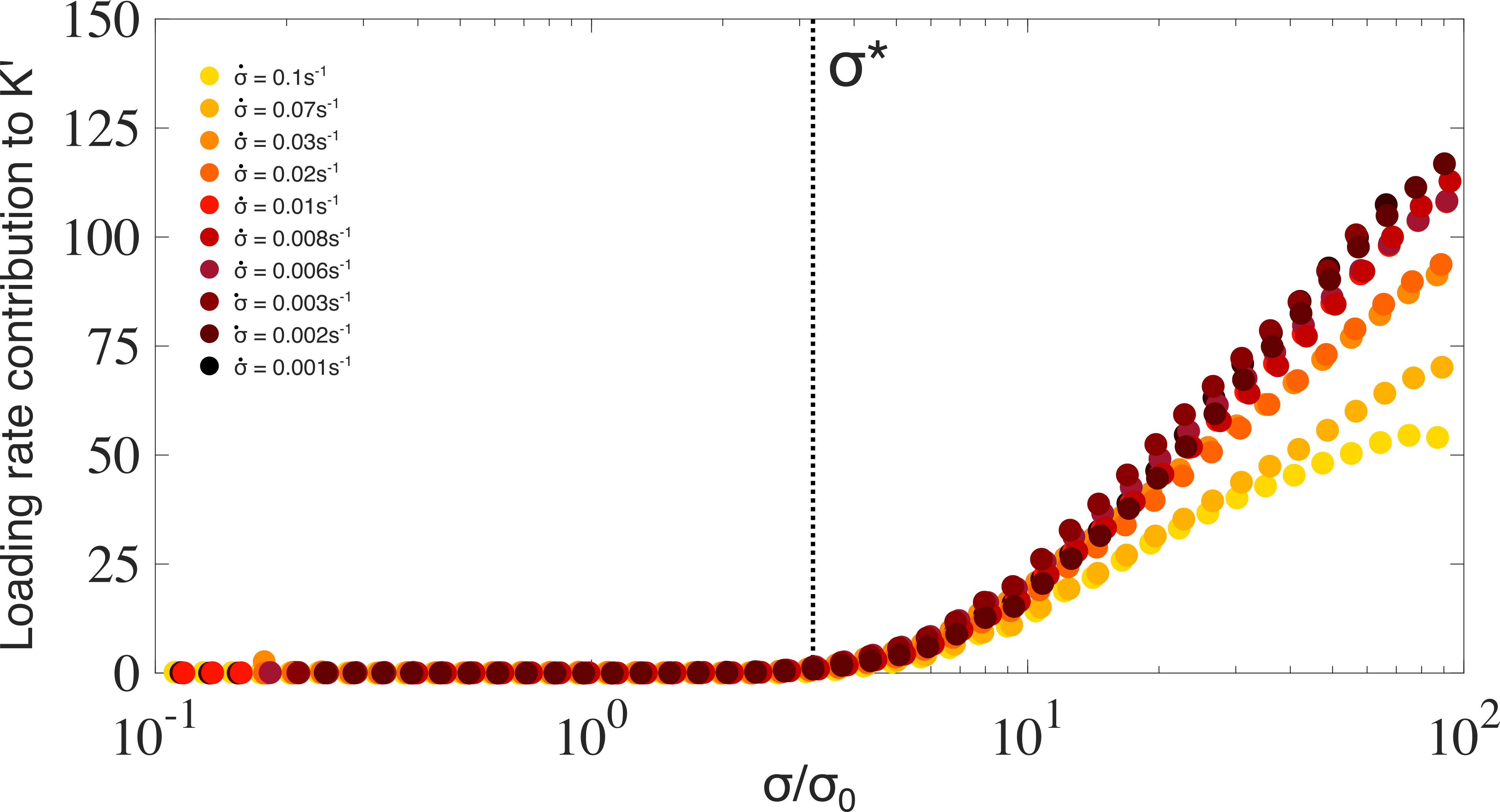 Figure 1
Figure 1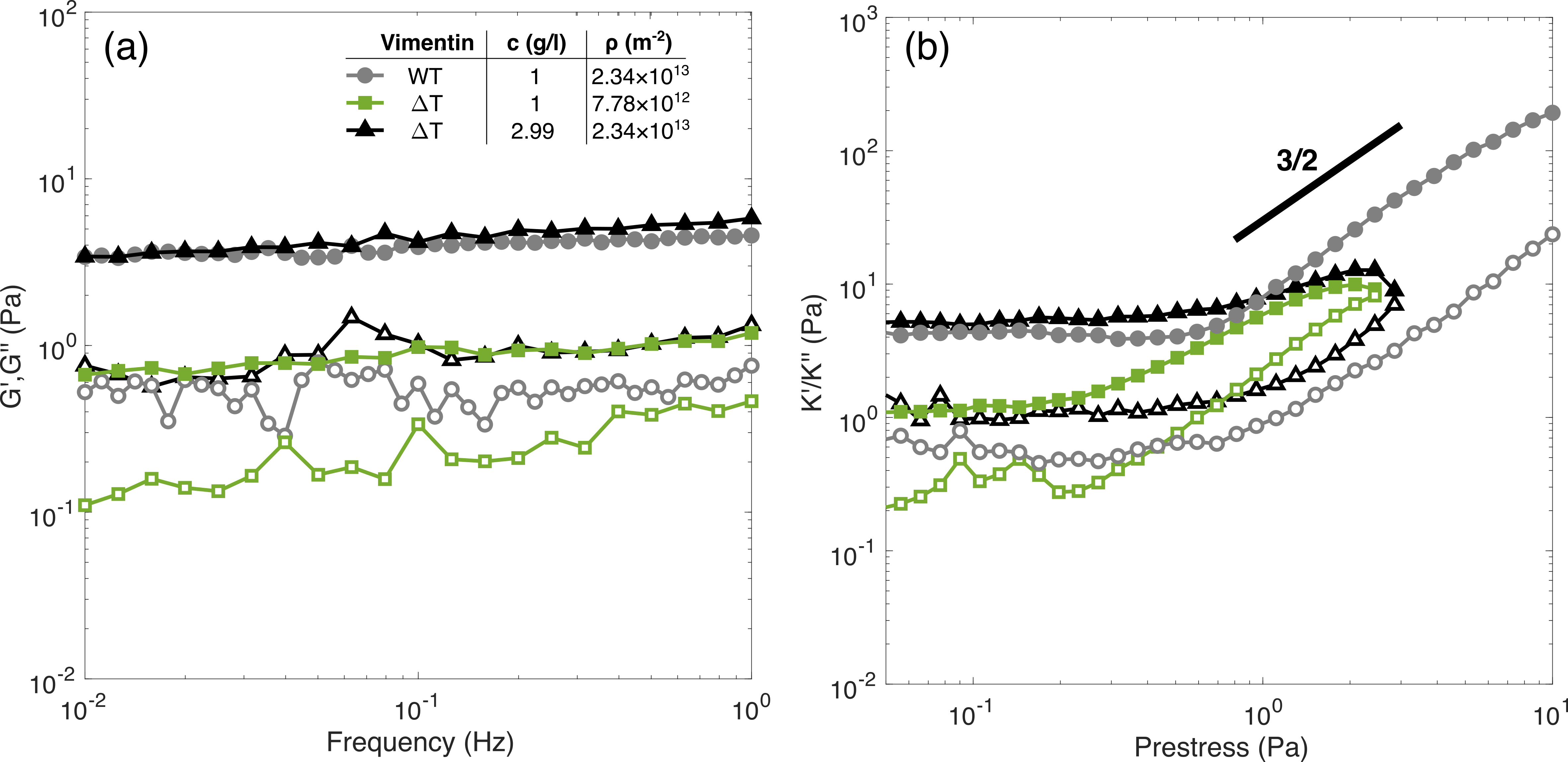 Figure 2
Figure 2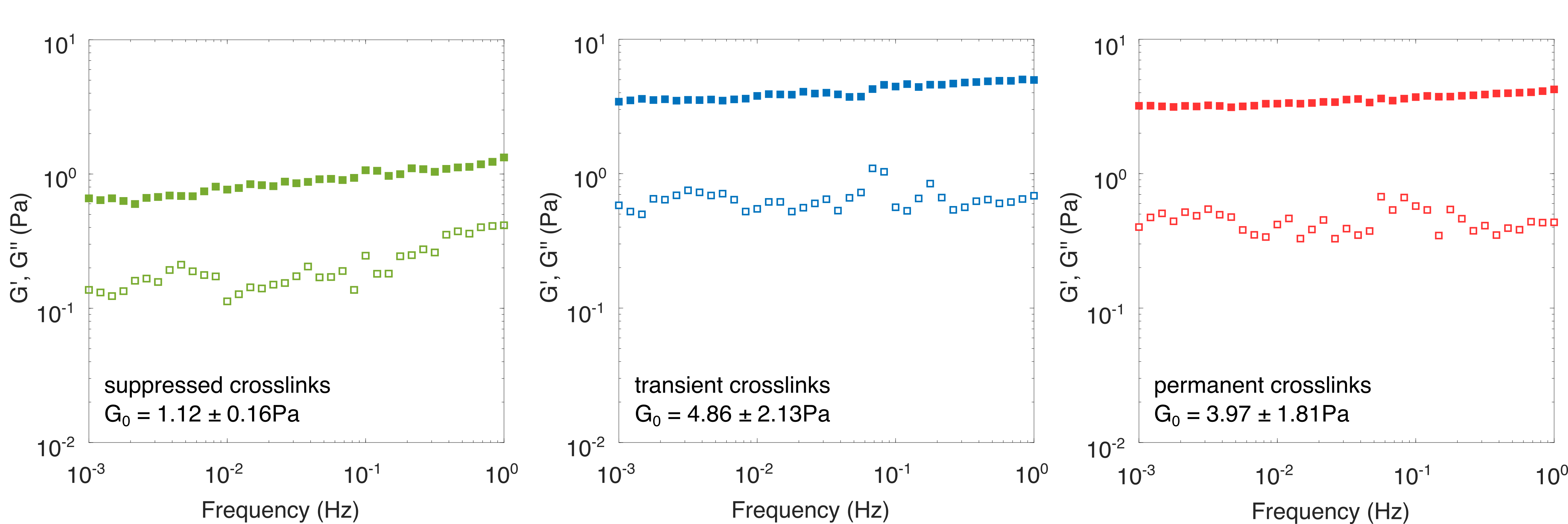 Figure 3
Figure 3 Figure 4
Figure 4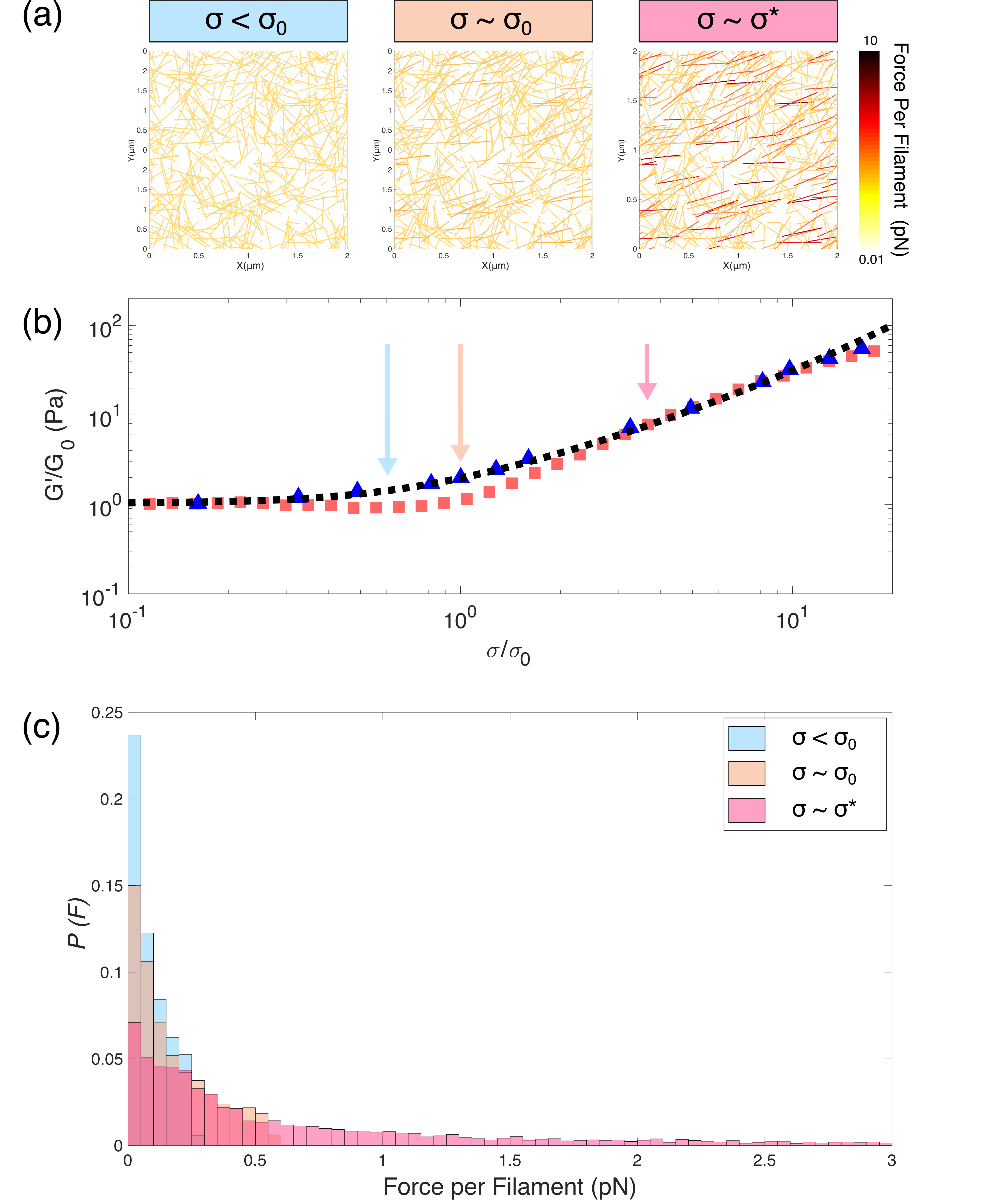 Figure 5
Figure 5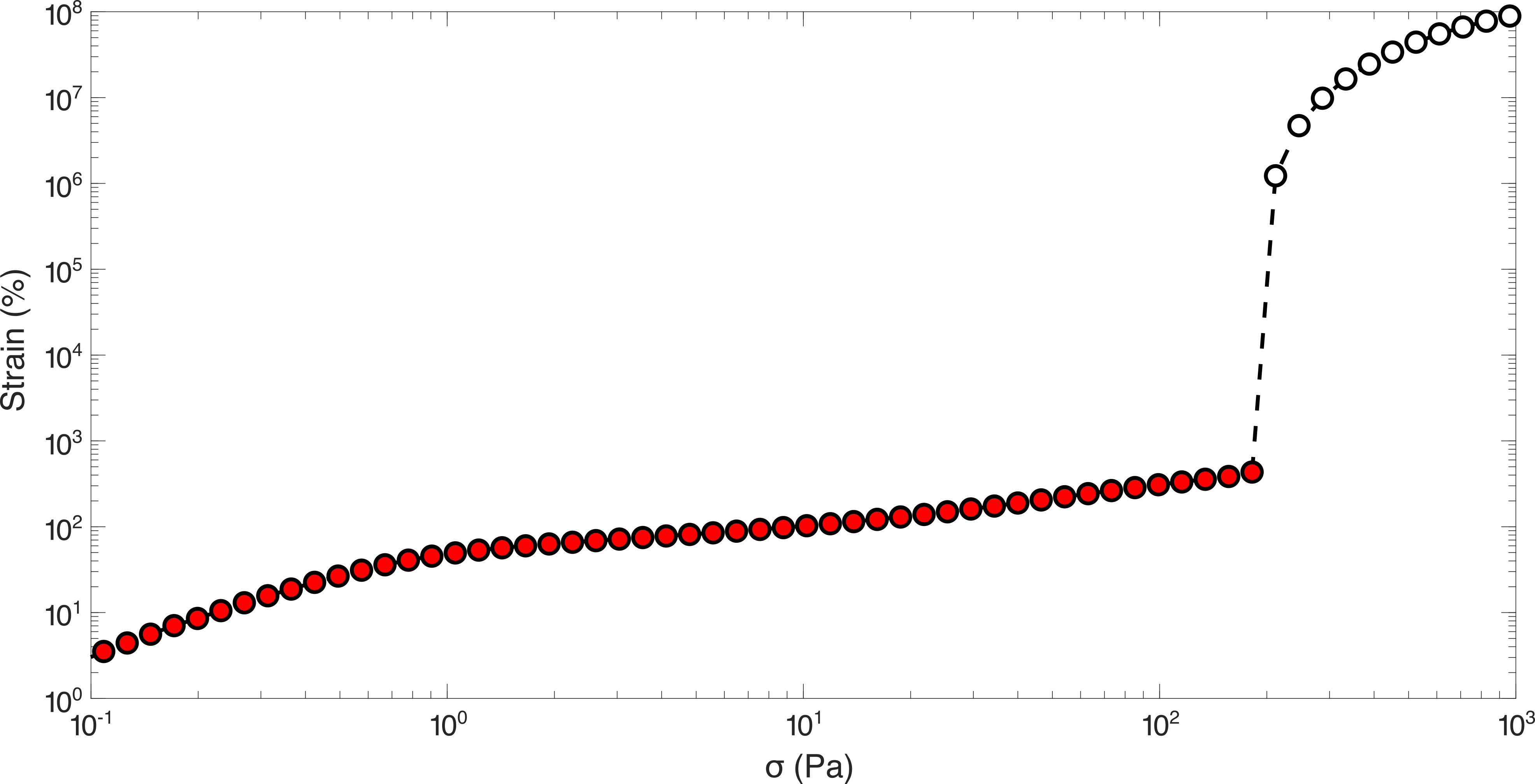 Figure 6
Figure 6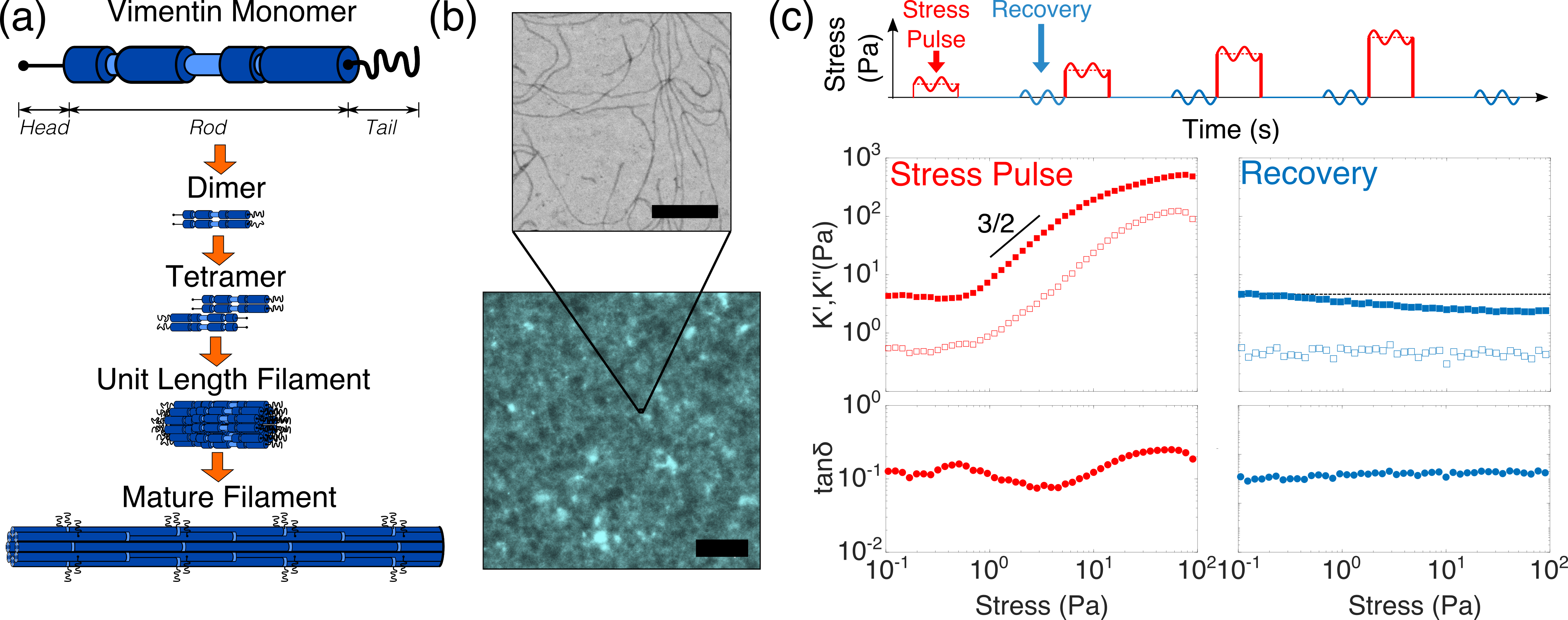 Figure 1
Figure 1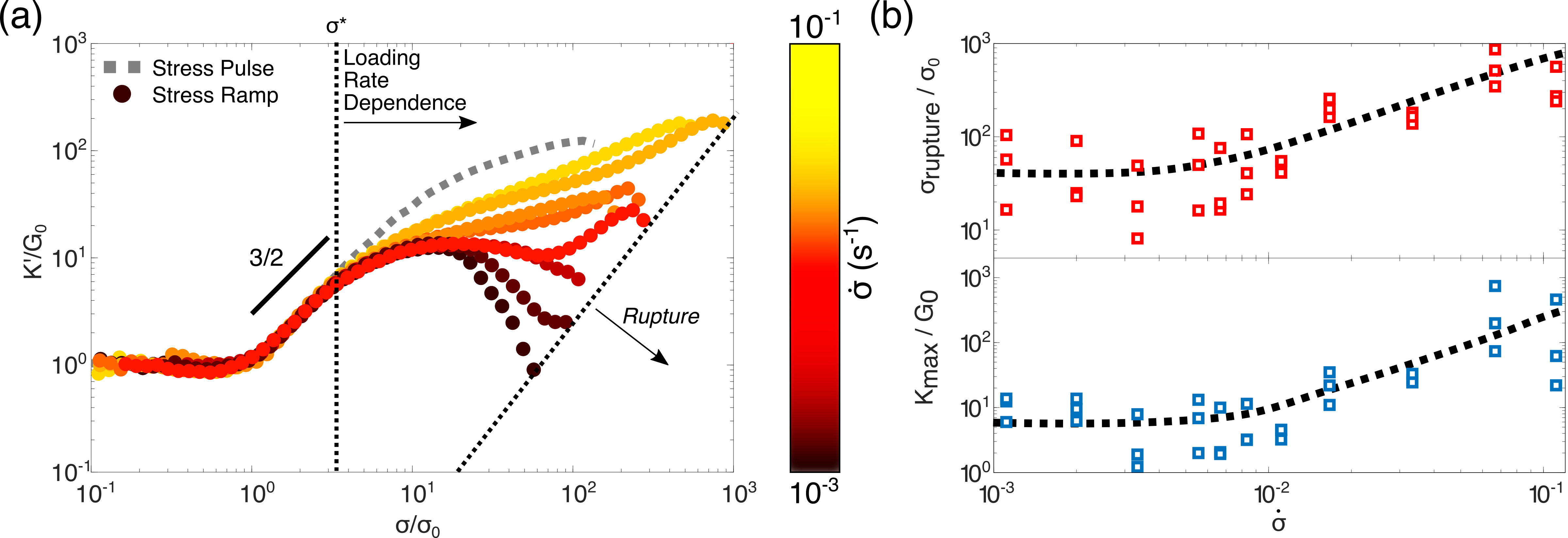 Figure 2
Figure 2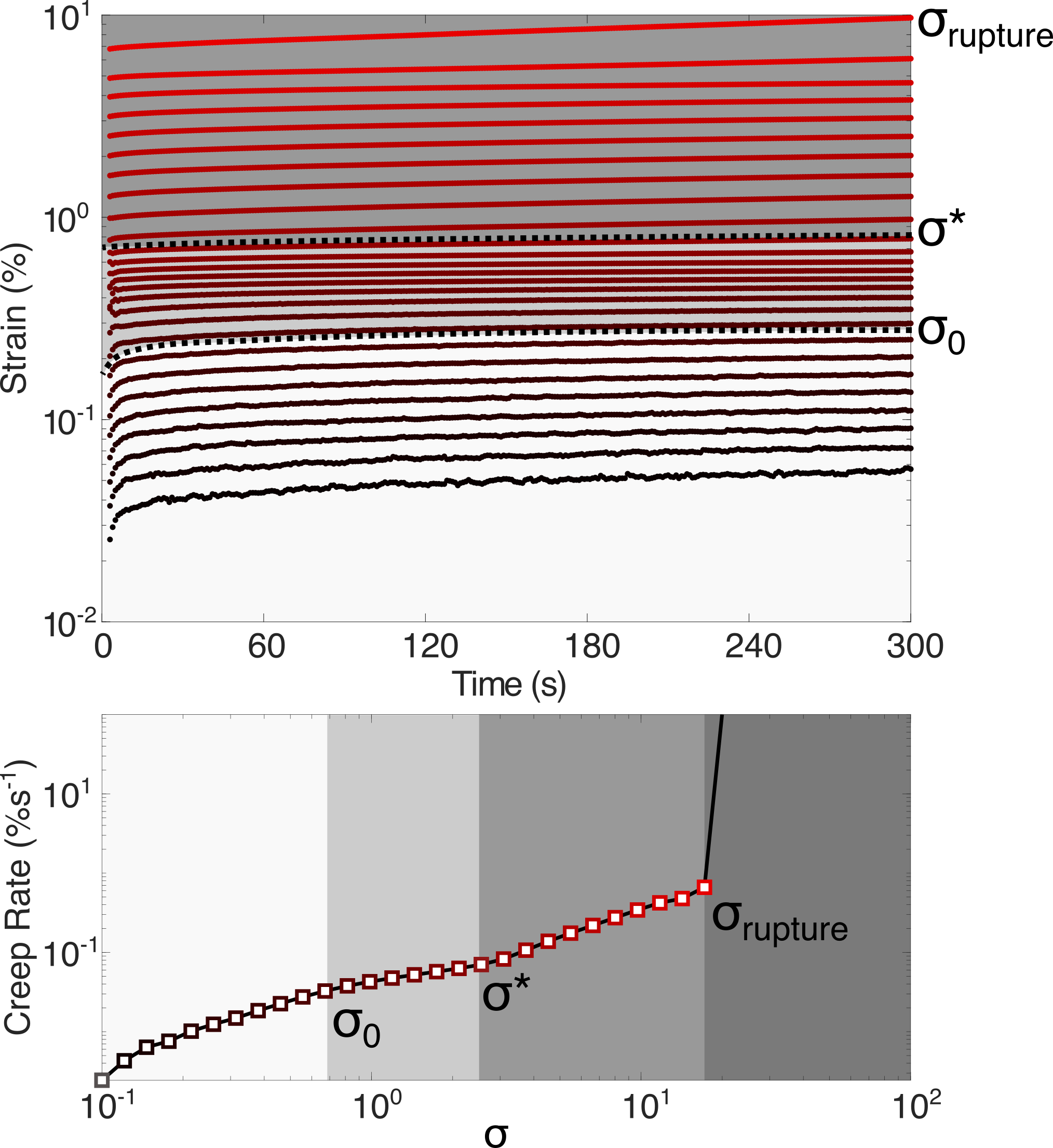 Figure 3
Figure 3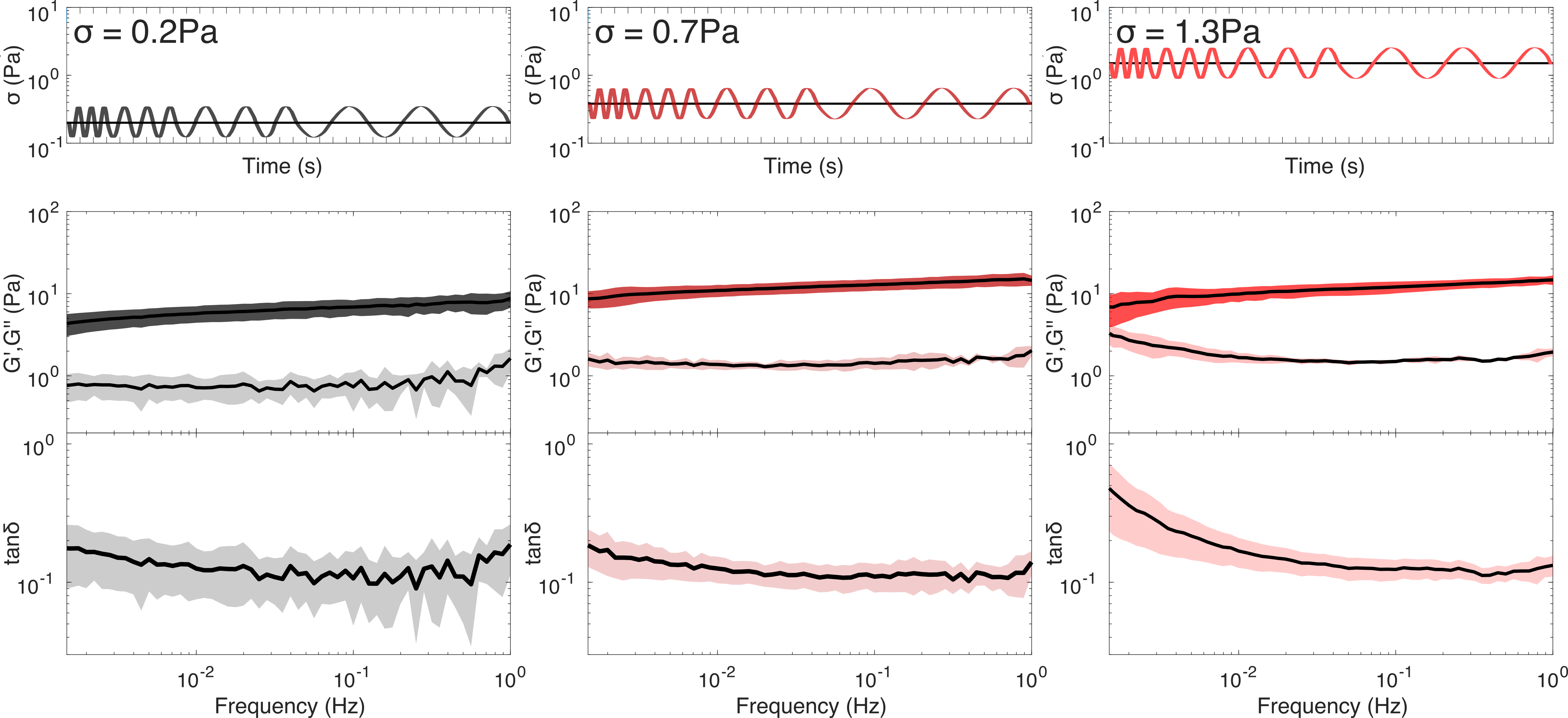 Figure 4
Figure 4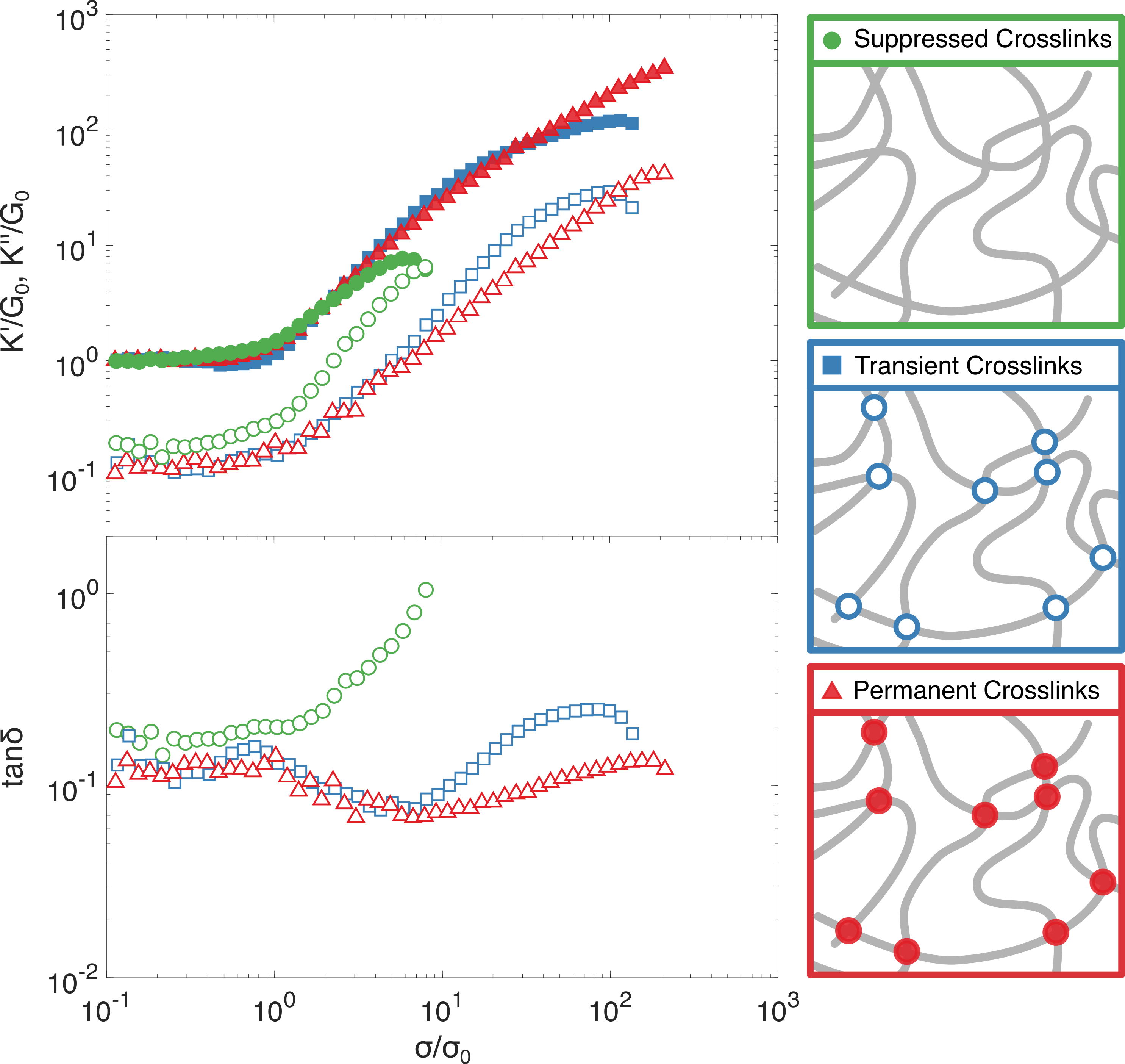 Figure 5
Figure 5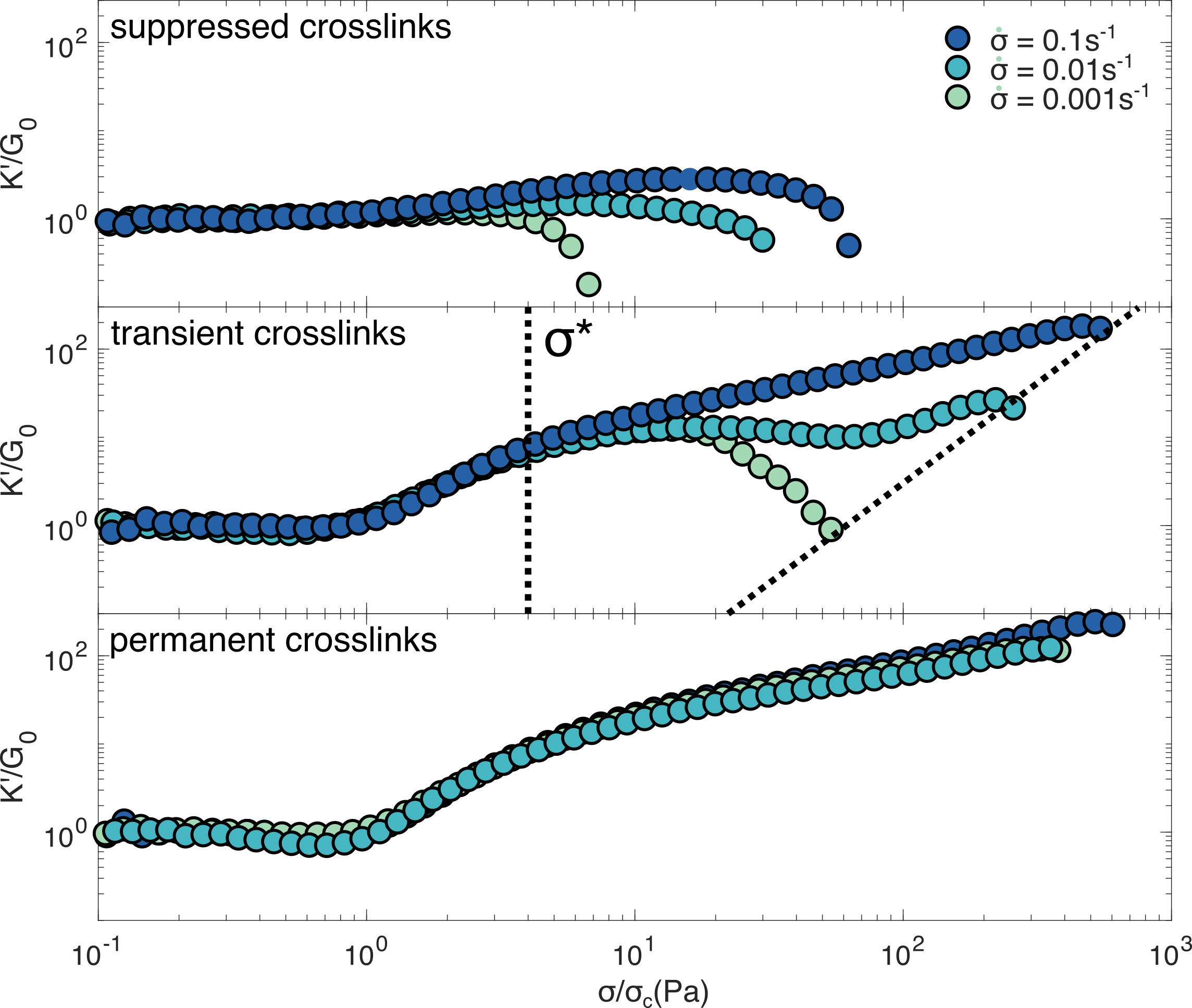 Figure 6
Figure 6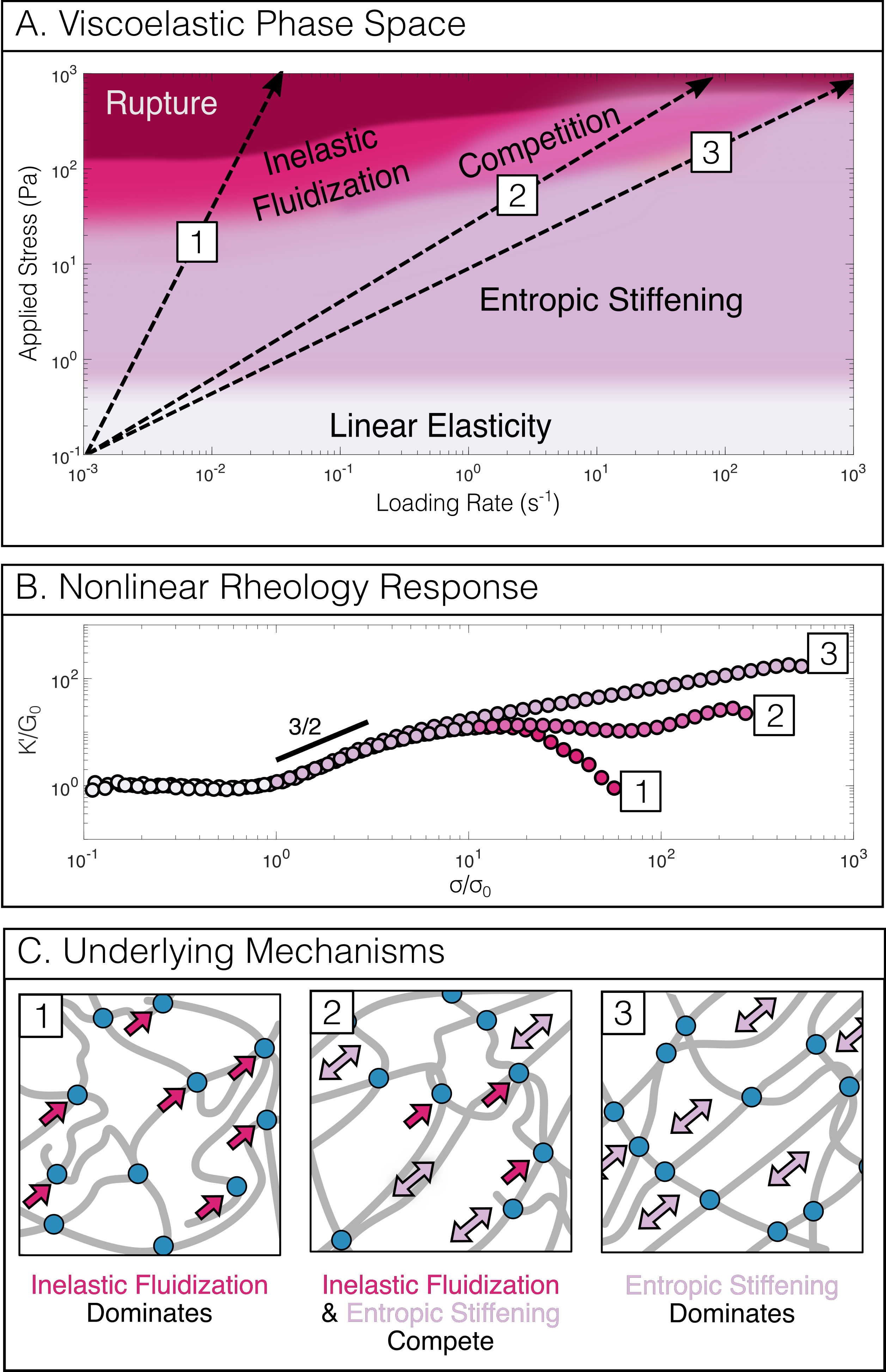 Figure 7
Figure 7Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Stiffening and Inelastic Fluidization in Vimentin Intermediate Filament Networks
Anders Aufderhorst-Roberts
Living Matter Department
AMOLF
1098 XG Amsterdam
The Netherlands
&Gijsje H. Koenderink
Living Matter Department
AMOLF
1098 XG Amsterdam
The Netherlands
[email protected] Present address:* School of Physics and Astronomy, University of Leeds, Leeds LS2 9JT, United Kingdom.*
Abstract
Intermediate filaments are cytoskeletal proteins that are key regulators of cell mechanics, a role which is intrinsically tied to their hierarchical structure and their unique ability to accommodate large axial strains. However, how the single-filament response to applied strains translates to networks remains unclear, particularly with regards to the crosslinking role played by the filaments’ disordered “tail” domains. Here we test the role of these noncovalent crosslinks in the nonlinear rheology of reconstituted networks of the intermediate filament protein vimentin, probing their stress- and rate- dependent mechanics. Similarly to previous studies we observe elastic stress-stiffening but unlike previous work we identify a characteristic yield stress , above which the networks exhibit rate-dependent softening of the network, referred to as inelastic fluidization. By investigating networks formed from tail-truncated vimentin, in which noncovalent crosslinking is suppressed, and glutaraldehyde-treated vimentin, in which crosslinking is made permanent, we show that rate-dependent inelastic fluidization is a direct consequence of vimentin’s transient crosslinking. Surprisingly, although the tail-tail crosslinks are individually weak, the effective timescale for stress relaxation of the network exceeds 1000s at . Vimentin networks can therefore maintain their integrity over a large range of strains (up to 1000) and loading rates (10*-3* to 10). Our results provide insight into how the hierarchical structure of vimentin networks contributes to the cell’s ability to be deformable yet strong.
00footnotetext: † Electronic Supplementary Information (ESI) available: Onset stress and linear modulus at all loading rates (S1), Representative stress vs. strain curve indicating rupture criteria (S2), loading rate dependent contributions to differential storage modulus (S3), affine entropic simulation data of vimentin networks (S4), frequency sweeps of vimentin networks with different crosslink modifications (S5), linear and nonlinear rheology of tailless vimentin (S6).
1 Introduction
The cells in our body have the remarkable ability to maintain but also adjust their structure in dynamic and stressful environments.11 This remarkable versatility is facilitated by the cytoskeleton, an intracellular filamentous network that spans the cell’s interior. Cells are able to stiffen in response to mechanical cues74, 64, 17, 41 but have also been shown to soften73, 43, 12 depending on the magnitude of applied stress,22 the loading rate 40 and the contributions of different cytoskeletal components 16 and the interactions between them.34
An integral subsystem of the cytoskeleton are the intermediate filaments, which form cell-spanning networks that are essential to mechanotransduction and provide resistance to external loads.26 Intermediate filament networks are very soft at small deformations but are able to stiffen substantially under stress29 and can withstand significantly higher axial strains than the other two cytoskeletal filaments, actin filaments and microtubules. 36 These unusual mechanical properties are thought to be related to the extensible -helical domains present in all intermediate filament monomers and the precise assembly sequence from monomers to filaments under physiological conditions72 (figure 1(a)). Their primary role as mechanoprotective elements is evidenced by the tendency of many cell types to increase their expression levels of intermediate filaments in response to stress19 and the numerous intermediate filament genetic mutations associated with diseases relating to cell fragility. 62 They are expressed in a cell- and organism- specific manner, suggesting their physiological function may relate directly to the local mechanical environment in different tissues.
Intermediate filament networks exhibit dynamic (dis)assembly and remodeling in the cell,67 but on a much larger timescale (minutes) compared to the turnover dynamics of actin filaments and microtubules (seconds), which enables them to act as long-lived structures that help maintain cell integrity28, 44 and cytoskeletal polarity39 even as cells perform dynamic processes such as migration.15, 21
How the interplay between dynamic remodelling of intermediate filaments and their nonlinear mechanics depends on the mechanical load and the loading rate remains an unresolved question. In cells, interactions between intermediate filaments are mediated by a variety of accessory proteins that crosslink the filaments to each other,76 to actin and microtubules,13 and to cell-matrix and cell-cell adhesions.66
Rheological studies on reconstituted networks of intermediate filaments have furthermore shown that they are inherently associative due to ionic interactions, 63 particularly between their highly charged carboxy-terminal tail domains which act as crosslinkers. 52 As a result of this crosslinking, intermediate filament networks exhibit nonlinear stress stiffening over several decades of applied stress.36 63, 69 The non-covalent nature of these interfilament interactions 37 presents a potential mechanism for dissipation, through a process known as inelastic fluidization.25 In contrast to entropic stiffening, which arises from the initial nonlinear stretch response of individual filaments,10 inelastic fluidization emerges from subsequent remodelling events such as unbinding and rebinding of the crosslinks between filaments leading to network softening. This behaviour was first observed in reconstituted actin networks,78, 48 in which transiently bound accessory proteins act as crosslinkers between filaments.49 Observing inelastic fluidization during mechanical loading is technically challenging because time-dependent and stress dependent mechanics are inherently convoluted in nonlinear rheology experiments.9 Moreover, it is challenging to discriminate inelastic effects at the network level from inelastic effects at the filament level. For intermediate filaments, these include rate-dependent extensibility5 and dissipation4 as measured by single-filament stretching experiments.
Here we investigate the consequences of noncovalent interactions for the nonlinear rheology of vimentin networks, using two complementary rheology protocols that allow us to deconvolve the time and stress-dependent mechanics. We are thus able to extract quantitative measures of the time scales and characteristic stress where inelastic fluidization occurs. We directly probe the contribution from noncovalent interactions between filaments by suppressing these interactions through genetic modification by removing the C-terminal tails and through chemical crosslinking. This comprehensive approach allows us to construct a phase space describing how the nonlinear response depends on loading rate and amplitude.
Methods
Protein Preparation
Human vimentin was expressed in Escherichia coli (strain TG1) and purified from inclusion bodies as previously described.32 A tailless mutant of human vimentin, with a truncation of the final 55 residues of the carboxy-terminal,31 was purified separately. Plasmid DNA for wild type and tailless vimentin were gifted by H. Herrmann, German Cancer Research Centre. Purified vimentin protein was stored at -80∘C in a storage buffer comprising 8 M urea, 5 mM Tris-HCl, pH 7.5, 1 mM dithiothreitol (DTT), 1 mM ethylenediaminetetraacetic acid (EDTA), 0.1 mM ethylene glycol bis(-aminoethyl ether)N,N’-tetraacetic acid (EGTA). The protein was dialyzed from 8M urea in steps of decreasing urea concentrations, (6M, 4M, 2M, 1M) by dilution with 5 mM Tris-HCl, pH 8.4, 1 mM EDTA, 0.1 mM EGTA, and 1 mM DTT, at room temperature with dialysis tubing with a molecular weight cut off of 12,000-14,000 as previously described33. Dialysis was continued overnight at 4∘C against a final buffer of 5 mM piperazine-N,N’-bis(2-ethanesulfonic acid) (PIPES), pH 7.0, 1 mM EGTA and 1 mM DTT. The final protein concentration was determined through measurement of the UV absorbance at 280nm using extinction coefficients of 22450M*-1cm-1* for wild type vimentin and 24870*-1cm-1* for tailless vimentin. The scattering contribution to the UV spectra was measured by fitting a Lambert-Beer law within the range 320-340nm and subtracting the result from the measured spectra.3 Dialysed protein was aliquotted, flash-frozen in liquid nitrogen and stored at -80∘C. To initiate filament assembly into networks we added a 10 concentrated polymerization buffer to thawed vimentin at 37∘C to obtain final concentrations of 25mM PIPES (pH 7.0), 100mM KCl, 4mM MgCl2 and 1mM EGTA.
Shear Rheology
All rheology measurements were performed with a stress-controlled rheometer (Kinexus Malvern Pro) equipped with a steel cone and plate (20mm diameter, 1∘ cone angle). Immediately after mixing with polymerization buffer the sample was transferred to the rheometer. Samples were left to polymerize for 60mins at 37∘C between the rheometer plates by which time the storage modulus G’ was observed to reach a steady state value. Mineral oil (Sigma Aldrich) was applied to the air-sample interface to prevent drying. Vimentin was polymerised at a concentration of 1mg/ml, unless otherwise stated. Assuming a mass per length of 6.65g/m,30 the mesh size of the network is estimated by the relation , where is the filament density expressed in terms of the total filament length per volume. A concentration of 1mg/ml of wild type vimentin with molecular weight 53.6kDa corresponds to = 206nm.
We term the two main nonlinear rheology protocols in this work the “stress pulse” and “stress ramp” protocols. In the stress pulse protocol, a steady prestress was applied for 20s, superimposed with a small amplitude oscillatory stress . The resulting oscillatory strain response was measured at a frequency of 0.5Hz. The oscillatory stress was fixed at of the steady prestress and the first 10s of oscillations were discarded to eliminate the possible influence of instrument inertial contributions. The stress was set to 0Pa for a period of 60s between successive measurements to probe network recovery following perturbation. The complex differential modulus in this approach, , is an instantaneous measurement9 and independent of inelastic fluidization over longer timescales. From the complex modulus, we derive the differential storage modulus, (K’) the differential loss modulus (K”) and the loss tangent (K”/K’). By contrast, in the stress ramp protocol,47, 71 no oscillations were superimposed, and the sample strain was measured while the steady stress was increased at a fixed logarithmic rate , defined in units of decades of applied stress per second. The resulting differential modulus K’ was calculated by applying a numerical derivative to the stress-strain curve. In the stress ramp protocol, K’ is dependent both on the elastic response of the constituent filaments and any inelastic fluidization, as demonstrated in earlier studies of entangled actin networks.9, 71
The time-dependence of network deformation over longer time scales was probed by a creep test, in which a constant shear stress was applied for 600 seconds and the resulting strain was measured. The creep rate was determined by performing a linear fit over the final 60s of the time dependent strain. The timescale for stress relaxation (where the elastic and viscous moduli cross-over) were probed by frequency-dependent oscillatory measurements over a frequency range of 10Hz, with a strain amplitude of 0.5. When probing the frequency response in the nonlinear regime, oscillations were superimposed on a steady stress through the same principles as outlined in the stress pulse protocol above. Repeated frequency sweeps were applied to each sample to verify that the samples were unaltered during the course of the measurement.
Fluorescence Imaging
Wild type vimentin was labelled through conjugation of its single cystein residue with Alexa Fluor 488 maleimide (Invitrogen), as previously described.77 Labelled and unlabelled vimentin were mixed in storage buffer before dialysis, with a fraction of labelled vimentin of approximately 5. Imaging was carried out using a Nikon A1 confocal microscope with a perfect focus system, a 100x/NA1.4 oil immersion objective, and a 100-mW 488 nm argon ion laser.
Scanning Transmission Electron Microscopy
Imaging was performed as previously described54 on a Verios 460 electron microscope (FEI) operating in bright field mode at an acceleration voltage of 20 kV. Vimentin filaments were assembled at a concentration of 0.01mg/ml for 1 hour and deposited on carbon-coated copper grids (Ted Pella). After 1 minute of deposition, the grids were rinsed with assembly buffer and adsorbed filaments were fixated by incubating with 0.1(w/v) glutaraldehyde dissolved in Milli-Q water (Merck) for 5 minutes. Before imaging, grids were rinsed 5 times with Milli-Q water and air-dried.
Network Modelling
In order to calculate the expected rheological response in the limit of affine (uniform) network deformation, we computed the stress-strain response of isotropic ensembles of non-interacting rigid rods. Isotropic ensembles of filaments were generated by a custom-written Matlab code. A total of 104 filaments of length were randomly deposited within a three-dimensional cube. The value of was inferred from measurements of the linear storage modulus using the relation:55 where is the Boltzmann constant, T is the experimental temperature of 310K and is the persistence length of vimentin (2.1m61). The length density of filaments in the simulation was selected to match the experimental length per volume . Each filament was assigned a random orientation and subjected to a homogeneous shear strain. The resulting filament tension was computed through an analytical expression for the force-extension relation of an inextensible semiflexible polymer.10
Results
We first compare the nonlinear viscoelastic response of vimentin networks through two rheology protocols. Figure 1(c) shows the results from the “stress pulse” protocol which probes the elasticity of the network in response to an instantaneous stress.23 As the stress is increased above a threshold stress , both the differential storage modulus K’ and loss modulus K” are observed to increase, indicating network stiffening. K’ increases with a power law of exponent 3/2, consistent with models of entropically-driven stiffening of semiflexible polymers and indicating that the network response inherits the force extension behaviour of its constituent filaments.55, 10 At higher stress, the stiffening exponent begins to level-off, indicating the enthalpic stretching of the individual filaments. 50, 52
The dissipation of stress in the vimentin network is quantified by the loss tangent, tan. The instantaneous nature of the stress pulse protocol means that dissipation has its origins in the viscous drag on the fluctuating filament segments.24 A slight increase in tan is observed immediately prior to the onset of stiffening, which we speculate may be due to the unbinding of weaker binding sites that have previously been reported to exist on vimentin’s rod domain63. In the stress regime where the network stiffens, tan decreases due to the damping of filament fluctuations.24 Only at high stress does tan begin to increase, indicating progressive crosslink unbinding and incipient network fracture. This leads to eventual network rupture, defined as tan, consistently measured as over 3 repeat experiments. We examine the plasticity of the network across these different regimes of applied stress by probing the mechanics following 60s of strain relaxation. A remarkably high degree of recovery of the original modulus of the virgin network is observed as, even immediately prior to the rupture of the network, G’ decreases no more than 2-fold in comparison to the original linear modulus. We note that this behaviour contrasts strongly with actin networks, which retain a strong “memory” of prior cycles of stress68 and strain.56
So far, we have examined only the instantaneous elasticity of the vimentin networks. To examine time-dependent mechanics we probe the same networks with the stress ramp protocol. Here, the measured mechanical response is the result of a convolution of two separate phenomena, the entropic stiffening previously shown in figure 1 and the inelastic fluidization of the vimentin network over time. Direct agreement between the protocols would therefore indicate the complete absence of inelastic fluidization.47, 9, 71 Figure 2(a) shows the nonlinear response from the stress ramp protocol over a range of loading rates of 10*-1*-10*-3s-1*, where the rate denotes decades of applied stress per second. At low applied stress, the initial stiffening as well as the linear modulus and onset stress (see figure S1) are all in close agreement with the entropic stiffening measured by the stress pulse protocol. By contrast, above a threshold stress the nonlinear response depends strongly on the loading rate: with large deviations from the stress pulse protocol observed at the slowest loading rates, and more subtle deviations seen at the fastest loading rates. As a consequence, both the degree of stiffening (figure 2 (b), bottom) and the rupture stress,(figure 2 (b), top) for convenience defined as the stress corresponding to a 5 orders of magnitude increase in strain (figure S2), increase substantially with loading rate.
The loading-rate dependent nonlinear response indicates a softening of the network which counteracts the entropic stiffening that is barely perceptible at the fastest loading rates () but clearly present at the slowest loading rates (). At intermediate loading rates (), we observe reentrant entropic stiffening, suggesting a sensitive interplay between entropic stiffening and softening. Remarkably, the onset of inelastic fluidization at is independent of loading rate (figure S3).
To independently verify the stress dependence of inelastic fluidization, we carry out a series of creep experiments, measuring the time-dependent strain response of the network at different levels of stress. Figure 3 shows the time dependent strain at increasing applied stress (top) and the associated creep rate (bottom). For the creep and creep rate both increase steadily with increasing stress, reaching a plateau at . At higher stress (), a subsequent increase in creep and creep rate is observed, confirming the presence of a inelastic fluidization at . Using the same definitions of the rupture stress as for the stress ramp protocol we find at applied strains of 1000, which corresponds approximately to the slowest loading rates in figure 2.
The hierarchical structure of vimentin networks presents a number of possible mechanisms by which inelastic fluidization could occur at . Firstly, it may be enthalpic, arising from the stretching of individual filaments beyond their equilibrium contour length via the unfolding of coiled-coil rod domains65, 5. To estimate the likelihood of this scenario, we calculate the expected forces exerted on individual filaments by modelling the network as an initially random ensemble of filaments that deforms affinely when a shear stress is applied, as shown in figure S4. The individual forces acting upon each filament at a given shear stress are determined by an analytical expression for the semi-flexible force-extension relation.10 The forces per filament are in the sub-piconewton range for . At a proportion of filaments () experience forces exceeding 1pN for , but these forces are much lower than those reported to initiate axial stretch of the filaments (of order 100s pN)5).
A more likely explanation is therefore that inelastic fluidization is caused by unbinding of crosslinks between the vimentin “tail” regions. The noncovalent nature of these crosslinks 37 suggests a mechanism for unbinding under tension and would also be consistent with the observed loading rate dependence because the bond fraction depends not only on the maximum load applied over the course of the experiment but also on total loading time.
To measure the stress-dependent crosslink unbinding time, we perform oscillatory frequency sweeps while simultaneously applying a constant stress, as shown in figure 4. We expect stress relaxation at frequencies below .8 At low prestress (0.2Pa), corresponding to the linear elastic regime previously identified in figure 1, both G’ and G” are nearly constant over all frequencies (figure 4, left) indicating minimal flow, which is consistent with the minimal creep observed at low stress (figure 3). Increasing the steady stress to a magnitude corresponding to (figure 4, centre) and beyond to (figure 4, right) reveals a convergence between G” and G’ at the lowest frequencies, indicating the onset of flow. Therefore, crosslink unbinding can be said to occur only over long timescales (exceeding the experimentally accessible time of 1000s) with increases in applied stress leading to faster unbinding kinetics, a phenomenon often referred to as “slip bond behaviour”.2
We have established that inelastic fluidization occurs above a threshold shear stress of (figure 2) corresponding to expected filament forces in the piconewton range (figure S4) at unbinding rates of order 10*-3*Hz (figure 4). To verify that inelastic fluidization is indeed caused by crosslink unbinding we seek to artificially modify the affinity and timescale of crosslinking. Firstly, we assemble the filaments in the presence of glutaraldehyde, which induces permanent bonding through fixation. 14 To minimize the risk of damaging45 or substantially stiffening the filaments through fixation46 we use low glutaraldehyde concentrations of 0.001(w/v), a 5 molar excess to vimentin. Fixed networks were found to have comparable loss and storage modulus to untreated networks, validating our approach (figure S5, middle and right panels). We also note previous reports that vimentin intermediate filaments retain their ability to axially stretch even when fixed with glutaraldehyde concentrations as high as 0.1(w/v)27. By contrast crosslinking is also suppressed by using a vimentin “tailless” mutant, which still leads to self-supporting networks (figure S5, left panel) a likely consequence of residual crosslinking provided by vimentin’s rod domain63 and possibly the effect of polymer entanglements.
The nonlinear mechanical response of these three crosslinking regimes (transient, permanent and suppressed crosslinking) using the stress pulse protocol are shown in figure 5. All three are observed to stiffen under applied stress, but the degree of stiffening and the rupture stress are strongly dependent on the nature of the crosslinking. Permanently crosslinked networks show the most pronounced stiffening and resilience to rupture while networks with suppressed crosslinking show minimal stiffening and lower rupture stresses. The permanent and transient networks exhibit a similar stiffening response up to stresses of approximately 10, the same point at which tan is observed to increase in transiently crosslinked networks (figure 5, bottom). A similar increase in tan is also observed when the crosslinking is suppressed but it occurs at significantly lower applied stress (), indicating a higher propensity for forced crosslink unbinding. Taken together these observations validate our hypothesis that force-dependent crosslink unbinding is the primary factor in determining inelastic fluidization.
Prior work has shown that tailless vimentin forms thicker and more irregular filaments compared to the full-length protein,31, 6 which would result in coarser networks at equivalent concentrations. To test the influence of this, we increase the concentration of tailless vimentin so that the filament contour length per unit volume matches that of the wild type network, using the previously reported data on the mass density of the filaments.31 For equivalent the linear mechanical response is identical between tailless and wild type vimentin networks (figure S5(a)) while the nonlinear mechanical response quantitatively resembles that of tailless vimentin at lower concentrations (figure S5 (b)). Importantly, also at equal fiber length density, the tailless vimentin stiffens much less than the full length protein, supporting our hypothesis that the tail domain has minimal influence on the linear modulus but is essential for robust strain stiffening.
Finally, we examine the loading rate dependent mechanics of the three different networks through the stress ramp protocol (figure 6). As expected, networks where crosslinking is suppressed exhibit minimal stress stiffening but pronounced softening at increasing loads, a phenomenon observed across all loading rates. By contrast, networks where crosslinking is permanent show high degrees of stiffening, with little sensitivity to loading rate. Thus, we can conclusively state that the loading rate dependent mechanics of vimentin networks is a direct consequence of their transient crosslinking.
Discussion
The overarching theme to our findings is that vimentin intermediate filament networks are not permanent structures but are dynamic as a consequence of the transient noncovalent interactions between the filaments. Similar characteristics have been observed in the other filamentous systems of the cytoskeleton, actin75 and microtubules51. In the case of these other cytoskeletal networks, inelastic fluidization appears to counteract their ability to withstand mechanical loads. By contrast, we show that vimentin networks are able to withstand much higher strains before rupture but also possess the ability to soften, counteracting the entropic stiffening.
Inelastic fluidization in intermediate filament networks is manifested by a characteristic yield stress , above which crosslink unbinding is enabled, and the network becomes highly sensitive to loading rate. This yield stress, defined by the onset of rate dependence in the stress pulse protocol (figure 2) lies comfortably within the range of contractile forces that are typically exerted by cells, of order 0.1-10Pa.38 Furthermore the networks do not permanently fracture since self-healing is observed upon removal of stress.
We estimate from assuming an affine network response that corresponds to forces exerted on individual filaments that are in the range of at most a few piconewtons (figure S4). Although rupture forces between tail domains have yet to be directly measured, the interaction potential between the disordered sidechains of neurofilaments, the intermediate filaments found in neuronal cells, is similarly low, of order of a few .1 These piconewton forces exerted on filaments are apparently sufficient to cause unbinding of the tail-mediated crosslinks. This implies that the tail-mediated crosslinks are significantly weaker than the crosslinks in actin networks provided by accessory crosslinker proteins, which exhibit typical rupture forces of order 10 pN.18 Nevertheless, reconstituted actin networks are able to withstand shear stresses that are about a factor of 10 lower than vimentin networks.40 We speculate that the differences between the strength of vimentin and actin networks may arise because binding between actin and its associated accessory proteins is defined locally by a single surface binding site while intermediate filament crosslinking is mediated by the collective interactions between their tail domains. Although these interactions are individually weaker, they may impart a stronger load resistance through a “strength in numbers” mechanism. Such a mechanism would suggest slower unbinding rates because unbinding would demand a collective rupture of all tail domains. Indeed we observe crosslink unbinding at particularly slow rates, (1000s at ), compared to the faster (10s) unbinding timescales of actin crosslinkers.60 A further possible factor in vimentin’s slow unbinding is that unbound tail domains will maintain a close proximity to each other if neighbouring tail domains remain bound. This would lead to an increased tendency for rebinding, in contrast to actin networks where accessory proteins are able to rapidly diffuse and bind elsewhere in the network. 59, 60 The slow dynamics we observe for vimentin filaments are consistent with the physiological role of intermediate filaments as maintainers of cell integrity70, 28, 44 and cytoskeletal polarity in migrating cells21, 39, 15 in direct contrast to actin’s role in “fast” cellular processes such as motility and shape change. 53
Based on our rheological data we propose a constitutive phase space for the nonlinear mechanics of intermediate filament networks (figure 7). This phase space reflects a balance between two phenomena: entropic stiffening at fast loading rates and low applied stress, and inelastic fluidization at slow loading rates and high applied stress. It has previously been proposed, that either one or the other phenomenon will dominate, depending on the relevant experimental time scale.78 The stiffening we observe is nearly instantaneous, occurring over the 20s timescale of the stress pulse protocol (figure 1), in direct contrast to the network’s slow inelastic fluidization. This large timescale separation enables the networks to exhibit a complex mechanical response in which entropic stiffening and inelastic fluidization act concurrently such that gradual crosslink unbinding can take place while filament segments between crosslinks continue to stiffen.
Conclusions
We have shown that intermediate filaments possess a complex mechanical response that depends both on the magnitude of applied stress and its timescale. By systematically probing the viscoelastic response as a function of load and loading rate, we highlight a number of previously unreported aspects of intermediate filament network mechanics. We show that the networks have a characteristic yield stress, above which the network softens in a loading-rate dependent manner. Above this yield stress, softening is highly sensitive to loading rate, a phenomenon that is shown to be a direct consequence of vimentin tail-mediated crosslinking. This crosslinking is transient but has a particularly slow unbinding rate, which we attribute to the collective binding of vimentin’s tail domains. The slow unbinding rate may explain why previous studies have indicated that intermediate filament crosslinking is permanent in nature.52
As well as being of fundamental interest, these slow unbinding rates may be of direct relevance to understanding force dissipation in the cytoskeleton. Previous work has attributed this dissipation to phenomena on the single filament level such as changes in protein secondary structure20, slippage of subunits58 and tail-domain unfolding.42 Our findings highlight that considerations at the network level must also be taken into account.
In the context of vimentin’s role in cell mechanics, it will be of particular interest to examine how crosslinking is regulated by the different concentrations of monovalent and divalent ions, which are known to influence the structure of the crosslink-mediating tail regions7 and their interaction strength.52 and by post-translational modifications such as phosphorylation of the tail domain.35 In addition it will be interesting to examine the interplay between noncovalent tail-mediated interactions and mobile crosslinkers by introducing reconstituted plectin into intermediate filament networks.57
Acknowledgments
We thank Chase Broedersz, Fred Mackintosh, Sarah Köster, Yuval Mulla and Bart Vos for many helpful discussions. We also thank Harald Herrmann for providing vimentin plasmids and for providing guidance with vimentin purification. We thank Marjolein Kuit-Vinkenoog and Jeffrey den Haan for help with protein purification and characterisation. We gratefully acknowledge funding from the Netherlands Organisation for Scientific Research (NWO), with a program grant from the Foundation for Fundamental Research on (FOM Program grant nr 143).
Supplementary Information
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 11 R. Beck, J. Deek, J. B. Jones, and C. R. Safinya. Gel-expanded to gel-condensed transition in neurofilament networks revealed by direct force measurements. Nature materials , 9(1):40, 2010.
- 22 G. I. Bell. Models for the specific adhesion of cells to cells. Science , 200(4342):618–627, 1978.
- 33 B. Birdsall, R. W. King, M. R. Wheeler, C. A. Lewis Jr, S. R. Goode, R. B. Dunlap, and G. C. Roberts. Correction for light absorption in fluorescence studies of protein-ligand interactions. Analytical biochemistry , 132(2):353–361, 1983.
- 44 J. Block, H. Witt, A. Candelli, J. C. Danes, E. J. Peterman, G. J. Wuite, A. Janshoff, and S. Köster. Viscoelastic properties of vimentin originate from nonequilibrium conformational changes. Science advances , 4(6):eaat 1161, 2018.
- 55 J. Block, H. Witt, A. Candelli, E. J. Peterman, G. J. Wuite, A. Janshoff, and S. Köster. Nonlinear loading-rate-dependent force response of individual vimentin intermediate filaments to applied strain. Physical review letters , 118(4):048101, 2017.
- 66 M. Brennich, U. Vainio, T. Wedig, S. Bauch, H. Herrmann, and S. Köster. Mutation-induced alterations of intra-filament subunit organization in vimentin filaments revealed by saxs. Soft matter , 2019.
- 77 M. E. Brennich, S. Bauch, U. Vainio, T. Wedig, H. Herrmann, and S. Köster. Impact of ion valency on the assembly of vimentin studied by quantitative small angle x-ray scattering. Soft Matter , 10(12):2059–2068, 2014.
- 88 C. P. Broedersz, M. Depken, N. Y. Yao, M. R. Pollak, D. A. Weitz, and F. C. Mac Kintosh. Cross-link-governed dynamics of biopolymer networks. Physical review letters , 105(23):238101, 2010.