Cortical Mirror-System Activation During Real-Life Game Playing: An Intracranial Electroencephalography (EEG) Study
Markus Kern, Johanna Ruescher, Andreas Schulze-Bonhage, Tonio Ball

TL;DR
This intracranial EEG study demonstrates that during real-life game playing, human mirror neuron system activity involves a broad cortical network, including premotor, Broca's, and temporo-parieto-occipital areas, during both movement observation and execution.
Contribution
First intracranial evidence showing that the human mirror system activates across a wide cortical network during natural, real-life motor behavior.
Findings
Mirror system activity involves premotor and motor cortex.
Cortical areas like Broca's and temporo-parieto-occipital junction are involved.
Spectral modulations are spatially correlated during observation and execution.
Abstract
Analogous to the mirror neuron system repeatedly described in monkeys as a possible substrate for imitation learning and/or action understanding, a neuronal execution/observation matching system (OEMS) is assumed in humans, but little is known to what extent this system is activated in non-experimental, real-life conditions. In the present case study, we investigated brain activity of this system during natural, non-experimental motor behavior as it occurred during playing of the board game "Malefiz". We compared spectral modulations of the high-gamma band related to ipsilateral reaching movement execution and observation of the same kind of movement using electrocorticography (ECoG) in one participant. Spatially coincident activity during both conditions execution and observation was recorded at electrode contacts over the premotor/primary motor cortex. The topography and amplitude of…
Click any figure to enlarge with its caption.
 Figure 1
Figure 1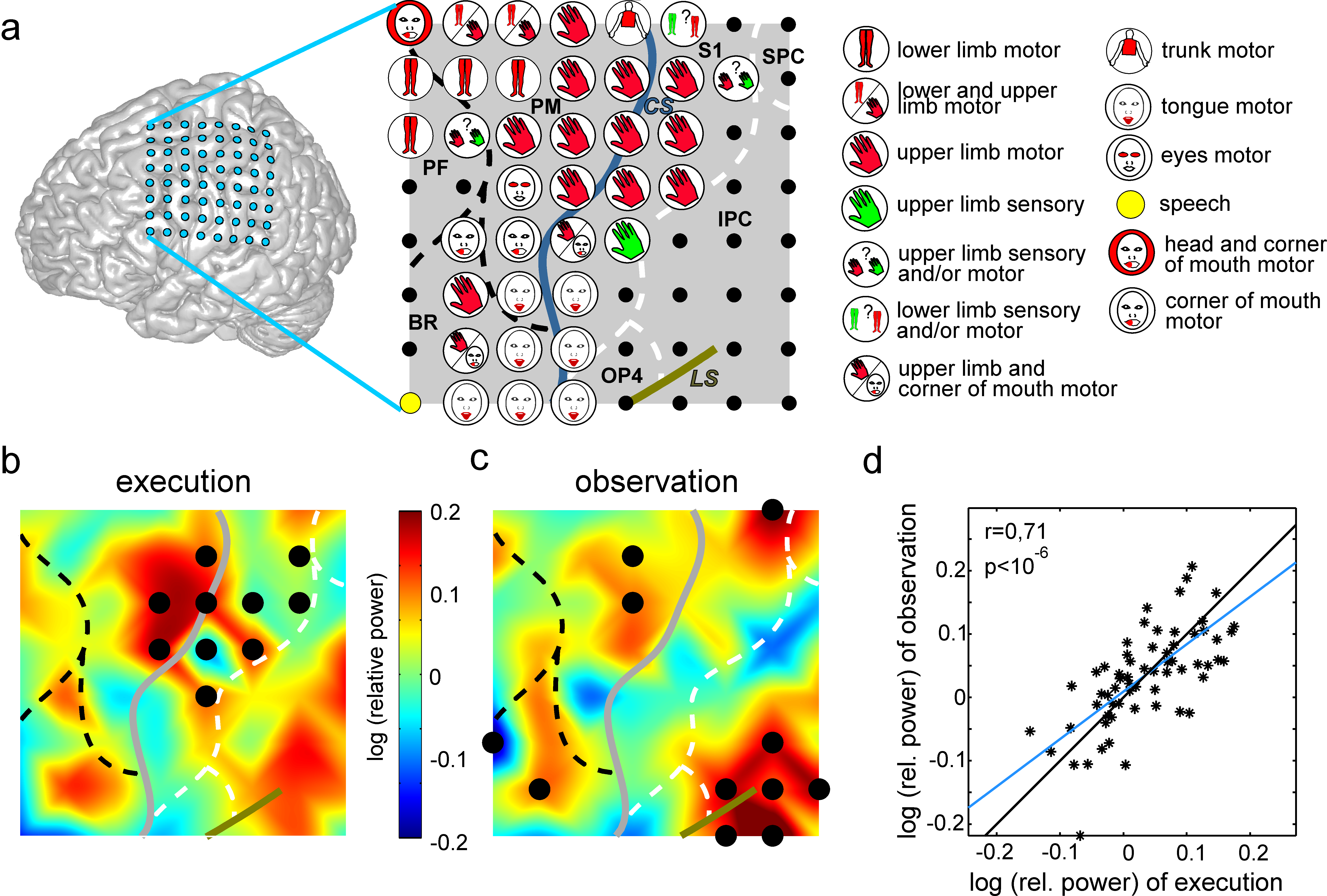 Figure 2
Figure 2Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Cortical Mirror-System Activation During Real-Life Game Playing:
An Intracranial Electroencephalography (EEG) Study
Markus Kern ([email protected])
BrainLinks-BrainTools Cluster of Excellence, University of Freiburg, Germany \ANDJohanna Ruescher ([email protected])
Faculty of Biology – University of Freiburg
79104, Freiburg, Germany \ANDAndreas Schulze-Bonhage ([email protected])
Epilepsy Center, Department of Neurosurgery, Medical Center — University of Freiburg,
79106, Freiburg, Germany \ANDTonio Ball ([email protected])
Medical AI Lab, University Medical Center Freiburg
79106, Freiburg, Germany
Abstract
**Analogous to the mirror neuron system repeatedly described in monkeys as a possible substrate for imitation learning and/or action understanding, a neuronal execution/observation matching system (OEMS) is assumed in humans, but little is known to what extent this system is activated in non-experimental, real-life conditions. In the present case study, we investigated brain activity of this system during natural, non-experimental motor behavior as it occurred during playing of the board game ”Malefiz”. We compared spectral modulations of the high-gamma band related to ipsilateral reaching movement execution and observation of the same kind of movement using electrocorticography (ECoG) in one participant. Spatially coincident activity during both conditions execution and observation was recorded at electrode contacts over the premotor/primary motor cortex. The topography and amplitude of the high-gamma modulations related to both, movement observation and execution were clearly spatially correlated over several fronto-parietal brain areas. Thus, our findings indicate that a network of cortical areas contributes to the human OEMS, beyond primary/premotor cortex including Broca’s area and the temporo-parieto-occipital junction area, in real-life conditions.
**
**Keywords: mirror system; ECoG; non-experimental; real life **
Introduction
Mirror neurons that respond both to (visually or auditory) movement observation and execution, have first been described in monkeys in area F5 of the ventral premotor cortex Pellegrino \BOthers. (\APACyear1992); Rizzolatti, Fadiga, Matelli\BCBL \BOthers. (\APACyear1996), in the inferior parietal cortex (IPC) Fogassi \BOthers. (\APACyear2005) and the intraparietal sulcus (IPS) Keysers (\APACyear2009). The analogous observation-execution matching system (OEMS) in humans, which is suggested to play a crucial role in imitation learning Iacoboni \BBA Wilson (\APACyear2006) and/or action understanding Rizzolatti, Fadiga, Gallese\BCBL \BBA Fogassi (\APACyear1996), is assumed to be more widely distributed over the human cortex than in monkeys Buccino \BOthers. (\APACyear2001); Keysers \BBA Gazzola (\APACyear2010). Indications of Human brain activity associated with the OEMS were observed in several cortical areas including the superior temporal sulcus (STS) Rizzolatti, Fadiga, Matelli\BCBL \BOthers. (\APACyear1996), Broca´s area (BR) Gazzola \BBA Keysers (\APACyear2009); Iacoboni \BOthers. (\APACyear1999); Rizzolatti, Fadiga, Matelli\BCBL \BOthers. (\APACyear1996), the IPC Gazzola \BBA Keysers (\APACyear2009); Newman-Norlund \BOthers. (\APACyear2007), the dorsal premotor cortex Gazzola \BBA Keysers (\APACyear2009), the ventral premotor cortex (vPM) Buccino \BOthers. (\APACyear2001); Dinstein \BOthers. (\APACyear2007); Filimon \BOthers. (\APACyear2015), the supplementary motor area (SMA) Gazzola \BBA Keysers (\APACyear2009); Hamzei \BOthers. (\APACyear2003); Iacoboni \BOthers. (\APACyear1999), the superior parietal cortex (SPC) Gazzola \BBA Keysers (\APACyear2009); Iacoboni \BOthers. (\APACyear1999), as well as the primary and secondary somatosensory cortex (S1/S2) Gazzola \BBA Keysers (\APACyear2009).
However, while these experimental studies clearly show the existence of an OEMS in the human brain, it is not clear whether the same kind of overlapping brain activities are also present during non-experimental, real-life conditions. Since striking differences in the firing behavior of cortical cells of the monkey brain were observed during natural movements compared to the instructed ones during experimental conditions Harrison \BBA Murphy (\APACyear2014), the investigation of human brain activity in real-life conditions is of interest.
Thus, in the present study we collected ECOG data during natural movement execution and observation, i.e., while the patient was engaged in the board game “Malefiz” with her life partner. This particular situation was chosen for due to the following reasons: i) the high degree of uniformity of arm movements in the observation and execution condition, ii) in the playing situation, the visitor was sitting close enough to the patients’ bed to be filmed by the digital video recorded for medical reasons, capturing only the direct vicinity of the hospital bed and iii) the patients’ attention during observation trials was specifically concentrated on the movements/actions of her game partner.
Methods
Data from a female, left handed 41 year-old patient were used, who was suffering from pharmacological resistant epilepsy and had a subdural 8*8 platinum electrode grid implanted over the left fronto-parietal cortex for pre-surgical evaluation. The patient agreed by informed consent to scientific usage of the recorded data and the study was approved by the local ethics committee. ECoG was recorded with a sampling rate of 1024 Hz. Monopolar electrical stimulation was conducted by physicians of the University Hospital Freiburg using an INOMED NS 60 stimulator (INOMED, Germany) (Fig. 2a). Details about the stimulation procedure are described in Ruescher \BOthers. (\APACyear2013). Anatomical assignment of electrodes to functional brain areas have been performed using the hierarchic anatomical assignment approach described in Pistohl \BOthers. (\APACyear2012).
We analyzed the ECoG data recorded while the patient played the game “Malefiz” with her life partner. In this game, both partners performed similar reaching movements to move their tokens on the game board. Due to handedness (left) and hemisphere of implantation (left), we analyzed ipsilateral reaching movements of the patient in the execution condition, consisting of 120 trials (Fig.1a). In the observation condition, we analyzed 87 trials of the left (14 trials) and right (73 trials) arm of the patient’s visitor (Fig.1b). Both persons were monitored by digital video recorded for medical reasons with a 25-Hz sampling rate and 640x480 pixels resolution. Trials were pre-selected by visual inspection of video recordings and confirmed by the analysis of the EMG recordings of the right and left deltoid muscles of the patient.
Event-related spectral power was calculated using a multi-taper spectral analysis method Percival \BBA Walden (\APACyear2006) with time windows of 200 ms, 20-ms time steps and 5 Slepian tapers. Since event-related HG power is proposed to be a suitable marker of local brain activity Crone \BOthers. (\APACyear1998), we analyzed modulations of HG band power relative to the power in baseline trials (resting condition, no movement execution or observation of movement, 21 trials).
The significance of HG power modulations was calculated using a paired two-sided Wilcoxon’s sign test. P-values were corrected by using the false discovery rate (FDR) approach Benjamini \BOthers. (\APACyear2001) with a q-level of . For a comparison of the overall fronto-parietal brain activity captured by grid electrodes during both conditions, we calculated Spearman’s rank-based correlations coefficient for HG power related between the execution and observation condition over the 64 grid electrode contacts.
Results
Both the execution and observation condition showed widespread high gamma (HG) band modulations in frontal and parietal regions (Fig. 2b,c). For the execution condition (Fig. 2b), significant increases of HG power were found over S1, PM, and IPC, while significant HG power decreases were found at two electrodes above the central sulcus and in the ventral S1. In the observation condition (Fig. 2c), significant HG power increases were found in PM, temporo-parieto-occipital junction area (TPO), dorsal S1 and BR.
On the single electrode level, significant responses overlapped in dorsal premotor cortex in a region showing hand motor responses during electrical stimulation mapping (Fig. 2a). Comparison of HG response topographies recorded over the whole electrode grid revealed significant positive correlations with r=0.71 and (Fig. 2d).
Discussion
In the present case study, we investigated the human OEMS by analyzing HG modulations in ECoG recordings of a female epilepsy patient while she was playing a board game with her partner. We found a significant increase of HG power at electrodes over PM during ipsilateral reaching movements as well as during the observation of unilateral reaching movements of the partner. This coincidental activity in PM is in line with results of recent fMRI studies investigating the OEMS Gazzola \BBA Keysers (\APACyear2009).
Overlapping activity was also recorded over Broca’s Area, the temporo-parietö-occipital (TPO) junction area and dorsal S1, but did not reach significance in the execution condition. This might partially be ascribed to the laterality of movement execution, since a movement-related increase in HG power is rather weak in the ipsilateral hemisphere compared to the contralateral side. However, BR and S1 have also been associated to the OEMS in previous studies Gazzola \BBA Keysers (\APACyear2009); Iacoboni \BOthers. (\APACyear1999); Rizzolatti, Fadiga, Matelli\BCBL \BOthers. (\APACyear1996).
However, BR and S1 have also been associated to the OEMS in previous studies Gazzola \BBA Keysers (\APACyear2009); Iacoboni \BOthers. (\APACyear1999); Rizzolatti, Fadiga, Matelli\BCBL \BOthers. (\APACyear1996). TPO has been found to be active during action observation Hamzei \BOthers. (\APACyear2003) and might thus play a role in the OEMS, too.
Additionally, we found significant coincident activity in S1, PM, prefrontal cortex and BR in another epilepsy patient not included in this study, related to contralateral finger movement execution (for more details, see P1 in Ball \BOthers. (\APACyear2008)), compared to the observation of her partner manipulating a Rubik’s cube.
Taken together, these findings support the suggestion, that the coincident activity we recorded over PM, BR, TPO and S1 during real-life conditions may be ascribed to the human OEMS. Interestingly, when comparing HG power at every channel of the fronto-parietal electrode grid, we found strong correlations between HG band modulations related to both conditions. First of all, this finding confirms the idea of multiple brain areas contributing to the human OEMS. Furthermore, the similarity of recorded signals during movement observation and execution might also be of interest for future research in the area of brain-computer interfaces, where it is crucial to reliably differentiate between these two conditions.
Acknowledgments
We would like to thank Simon Contzen for his help in preparing this manuscript. This study was (partly) funded by the WISPER Project, Baden-Württemberg Stiftung.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ball \B Others . ( \APA Cyear 2008) \APA Cinsertmetastar ball_movement_2008 {APA Crefauthors} Ball, T., Demandt, E., Mutschler, I., Neitzel, E., Mehring, C., Vogt, K. \BDBL Schulze-Bonhage, A. \APA Cref Year Month Day 2008 \APA Cmonth 06. \BBOQ \APA Crefatitle Movement related activity in the high gamma range of the human EEG Movement related activity in the high gamma range of the human EEG. \BBCQ \APA Cjournal Vol Num Pages Neuro Image 412302–310. \Print Back Refs \Current Bib
- 2Benjamini \B Others . ( \APA Cyear 2001) \APA Cinsertmetastar benjamini_controlling_2001 {APA Crefauthors} Benjamini, Y., Drai, D., Elmer, G., Kafkafi, N. \BCBL \BBA Golani, I. \APA Cref Year Month Day 2001 \APA Cmonth 11. \BBOQ \APA Crefatitle Controlling the false discovery rate in behavior genetics research Controlling the false discovery rate in behavior genetics research. \BBCQ \APA Cjournal Vol Num Pages Behavioural Brain Research 1251-2279–284. \Print Back Refs \Current Bib
- 3Buccino \B Others . ( \APA Cyear 2001) \APA Cinsertmetastar buccino_action_2001 {APA Crefauthors} Buccino, G., Binkofski, F., Fink, G \BPBI R., Fadiga, L., Fogassi, L., Gallese, V. \BDBL Freund, H \BHBI J. \APA Cref Year Month Day 2001 \APA Cmonth 01. \BBOQ \APA Crefatitle Action observation activates premotor and parietal areas in a somatotopic manner: an f MRI study Action observation activates premotor and parietal areas in a somatotopic manner: an f MRI study. \BBCQ \APA Cjournal Vol Num P
- 4Crone \B Others . ( \APA Cyear 1998) \APA Cinsertmetastar crone_functional_1998 {APA Crefauthors} Crone, N \BPBI E., Miglioretti, D \BPBI L., Gordon, B. \BCBL \BBA Lesser, R \BPBI P. \APA Cref Year Month Day 1998 \APA Cmonth 12. \BBOQ \APA Crefatitle Functional mapping of human sensorimotor cortex with electrocorticographic spectral analysis. II. Event-related synchronization in the gamma band. Functional mapping of human sensorimotor cortex with electrocorticographic spectral analysis. II
- 5Dinstein \B Others . ( \APA Cyear 2007) \APA Cinsertmetastar dinstein_brain_2007 {APA Crefauthors} Dinstein, I., Hasson, U., Rubin, N. \BCBL \BBA Heeger, D \BPBI J. \APA Cref Year Month Day 2007 \APA Cmonth 09. \BBOQ \APA Crefatitle Brain Areas Selective for Both Observed and Executed Movements Brain Areas Selective for Both Observed and Executed Movements. \BBCQ \APA Cjournal Vol Num Pages Journal of Neurophysiology 9831415–1427. \Print Back Refs \Current Bib
- 6Filimon \B Others . ( \APA Cyear 2015) \APA Cinsertmetastar filimon_observed_2015 {APA Crefauthors} Filimon, F., Rieth, C \BPBI A., Sereno, M \BPBI I. \BCBL \BBA Cottrell, G \BPBI W. \APA Cref Year Month Day 2015 \APA Cmonth 09. \BBOQ \APA Crefatitle Observed, Executed, and Imagined Action Representations can be Decoded From Ventral and Dorsal Areas Observed, Executed, and Imagined Action Representations can be Decoded From Ventral and Dorsal Areas. \BBCQ \APA Cjournal Vol Num Pages Cereb
- 7Fogassi \B Others . ( \APA Cyear 2005) \APA Cinsertmetastar fogassi_parietal_2005 {APA Crefauthors} Fogassi, L., Ferrari, P \BPBI F., Gesierich, B., Rozzi, S., Chersi, F. \BCBL \BBA Rizzolatti, G. \APA Cref Year Month Day 2005 \APA Cmonth 04. \BBOQ \APA Crefatitle Parietal Lobe: From Action Organization to Intention Understanding Parietal Lobe: From Action Organization to Intention Understanding. \BBCQ \APA Cjournal Vol Num Pages Science 3085722662–667. \Print Back Refs \Current Bib
- 8Gazzola \BBA Keysers ( \APA Cyear 2009) \APA Cinsertmetastar gazzola_observation_2009 {APA Crefauthors} Gazzola, V. \BCBT \BBA Keysers, C. \APA Cref Year Month Day 2009 \APA Cmonth 06. \BBOQ \APA Crefatitle The Observation and Execution of Actions Share Motor and Somatosensory Voxels in all Tested Subjects: Single-Subject Analyses of Unsmoothed f MRI Data The Observation and Execution of Actions Share Motor and Somatosensory Voxels in all Tested Subjects: Single-Subject Analyses of Unsmoo