Shape Transformations of Vesicles induced by Swim Pressure
Yao Li, Pieter Rein ten Wolde

TL;DR
This paper investigates how active forces from Brownian particles inside vesicles induce diverse shape transformations, revealing novel phase transitions and dynamic behaviors through combined theoretical and simulation approaches.
Contribution
It introduces a comprehensive study of active force effects on vesicle shapes, uncovering new phase transitions and oscillatory behaviors not seen in passive systems.
Findings
Discontinuous spherical to prolate transition at low swim pressure
Stochastic oscillations at high swim pressure
Active forces induce novel shape dynamics in vesicles
Abstract
While the behavior of vesicles in thermodynamic equilibrium has been studied extensively, how active forces control vesicle shape transformations is not understood. Here, we combine theory and simulations to study the shape behavior of vesicles containing active Brownian particles. We show that the combination of active forces, dimensionality and membrane bending free energy creates a plethora of novel phase transitions. At low swim pressure, the vesicle exhibits a discontinuous transition from a spherical to a prolate shape, which has no counterpart in two dimensions. At high swim pressure it exhibits stochastic spatio-temporal oscillations. Our work helps to understand and control the shape dynamics of membranes in active-matter systems.
Click any figure to enlarge with its caption.
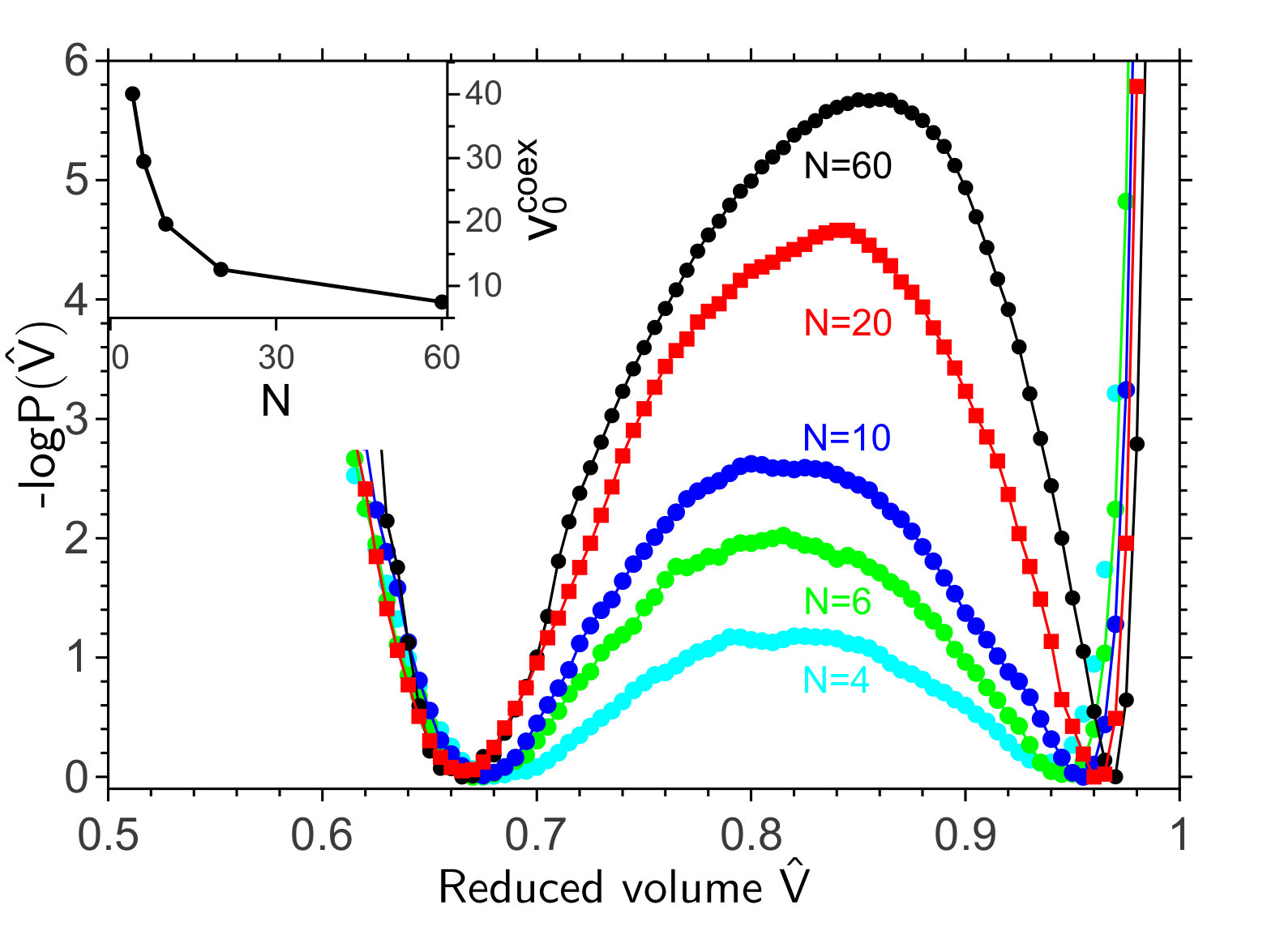 Figure 1
Figure 1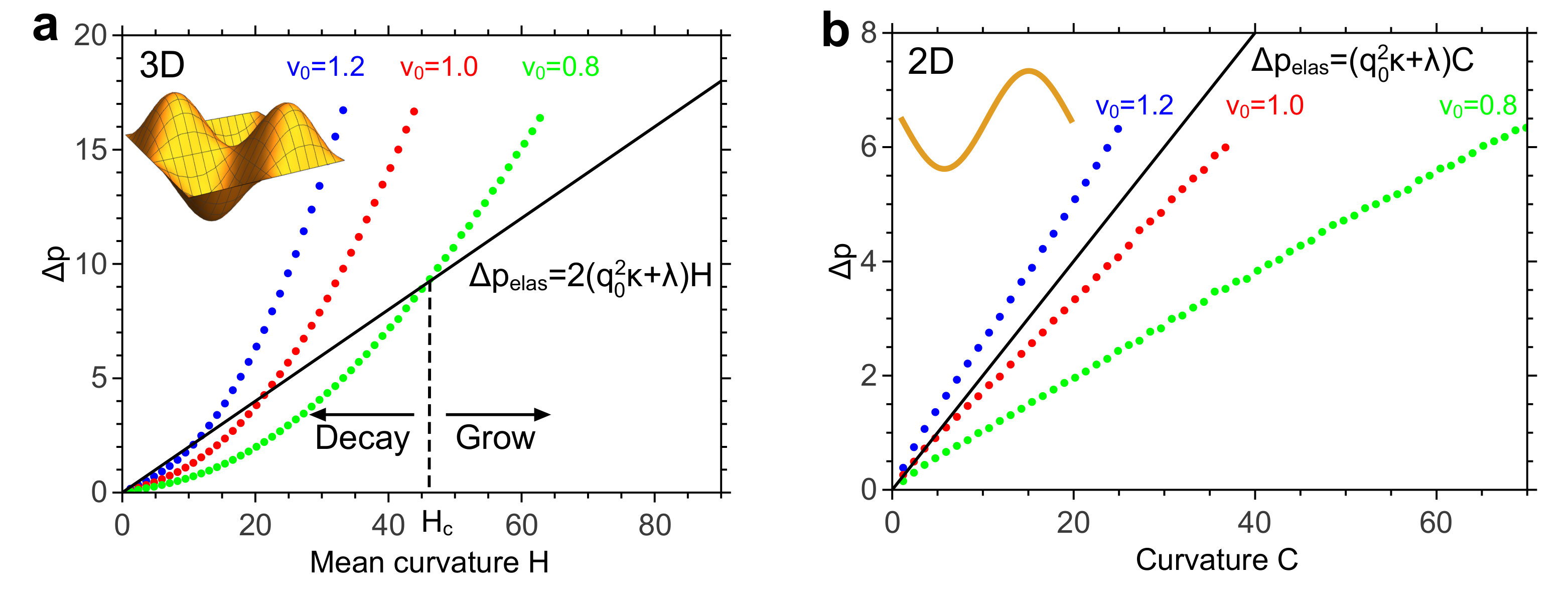 Figure 2
Figure 2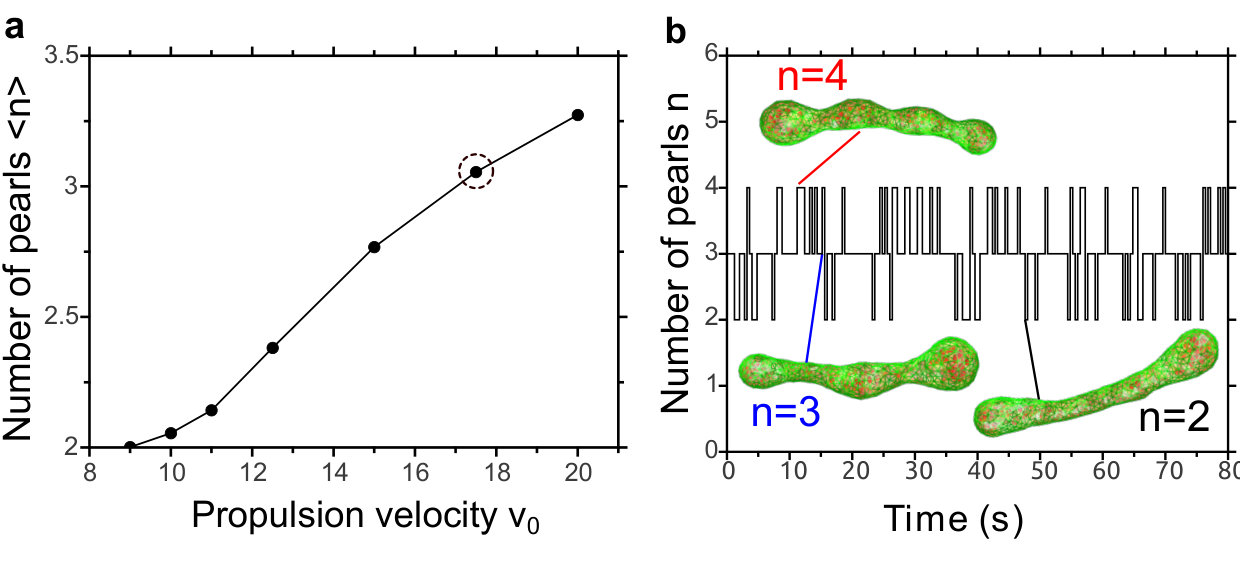 Figure 3
Figure 3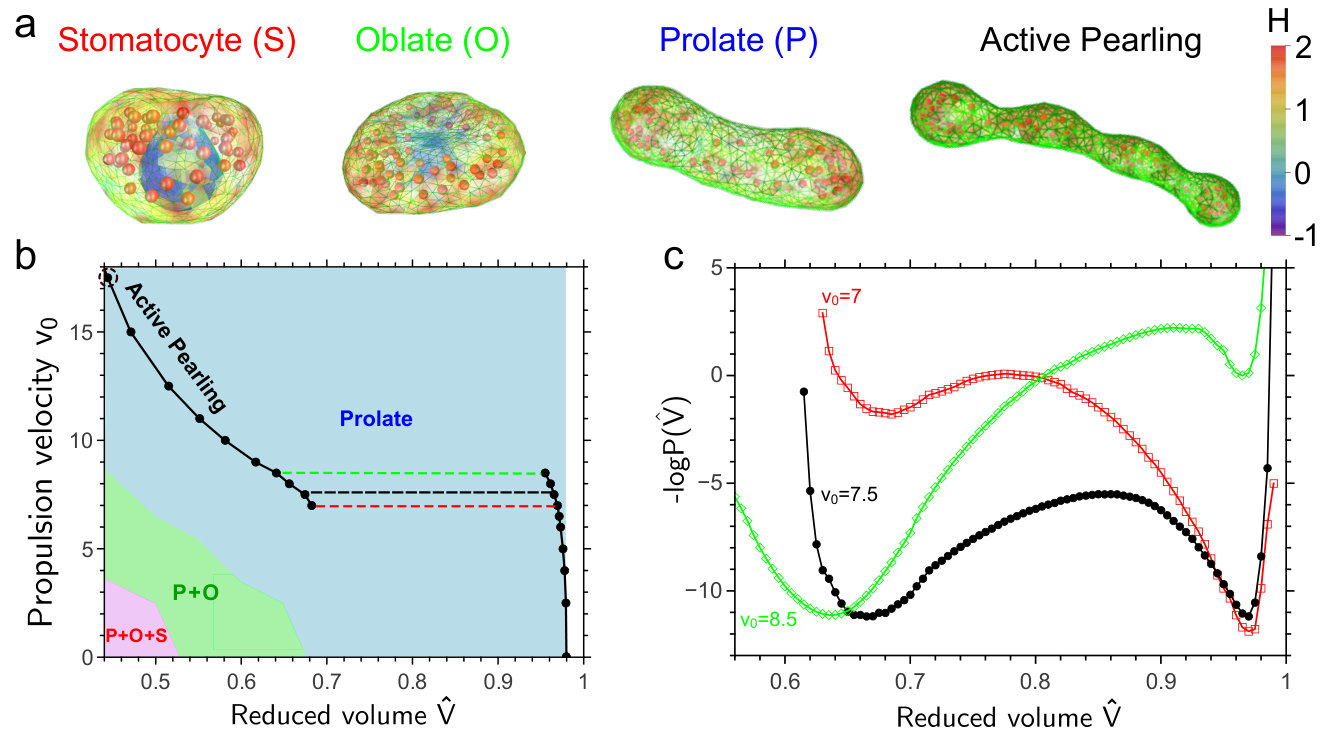 Figure 4
Figure 4Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Shape Transformations of Vesicles induced by Swim Pressure
Yao Li, Pieter Rein ten Wolde
AMOLF, Science park 104, 1098 XG Amsterdam, The Netherlands
Abstract
While the behavior of vesicles in thermodynamic equilibrium has been studied extensively, how active forces control vesicle shape transformations is not understood. Here, we combine theory and simulations to study the shape behavior of vesicles containing active Brownian particles. We show that the combination of active forces, dimensionality and membrane bending free energy creates a plethora of novel phase transitions. At low swim pressure, the vesicle exhibits a discontinuous transition from a spherical to a prolate shape, which has no counterpart in two dimensions. At high swim pressure it exhibits stochastic spatio-temporal oscillations. Our work helps to understand and control the shape dynamics of membranes in active-matter systems.
Active matter systems containing self-propelled units are not only omnipresent in the natural world Cavagna et al. (2017); Kearns (2010); Mora et al. (2016), but are also increasingly made synthetically Howse et al. (2007); Jiang et al. (2010); Wilson et al. (2012); Palacci et al. (2013); Bricard et al. (2013); Needleman and Dogic (2017). These systems are inherently out of thermodynamic equilibrium and show behavior that cannot be observed in equilibrium systems, such as spontaneous flow Voituriez et al. (2006); Galajda et al. (2007); Wan et al. (2008); Doostmohammadi et al. (2018), athermal phase separation Tailleur and Cates (2008); Redner et al. (2013); Buttinoni et al. (2013) or oscillations Keber et al. (2014). A key quantity in these active systems is the swim pressure Takatori et al. (2014); Yang et al. (2014); Solon et al. (2015a), which in contrast to the thermodynamic pressure depends on the shape of the boundary Solon et al. (2015b); Nikola et al. (2016); Junot et al. (2017). In both natural and synthetic systems, boundaries are often formed by soft lipid bilayers, which can change shape under the influence of pressure. Yet, the feedback between swim pressure and membrane shape dynamics is not understood.
Here we study shape transformations of vesicles containing active particles. Passive vesicles in thermodynamic equilibrium exhibit a variety of shapes, including prolate, oblate, and chain of “pearls” Seifert (1997); Yu and Granick (2009). These shapes are governed by the Helfrich bending free energy Helfrich (1973), the osmotic pressure, and, if applicable, the adsorption of particles onto the membrane Yu and Granick (2009). How active forces affect vesicle shape is, however, a wide open question Mietke et al. (2019). While the osmotic pressure controls vesicle shape only indirectly by setting the volume Seifert (1997), the swim pressure as generated by active Brownian particles Takatori et al. (2014); Yang et al. (2014); Solon et al. (2015a) also depends on the shape of the membrane Solon et al. (2015b); Nikola et al. (2016); Junot et al. (2017) Computational studies on active particles confined within a semi-flexible ring polymer in 2D show that active forces can change the shape of the polymer from circular to elliptical Tian et al. (2015); Paoluzzi et al. (2016), suggesting that the swim pressure can, unlike the osmotic pressure, also directly control vesicle shape. Yet how the swim pressure interacts with membrane elasticity to control shape, and how this interaction depends on the dimensionality of the system, are not understood. Here we show that this interaction is fundamentally different in 2D and 3D and that as a result 3D vesicles exhibit shape transformations not observed in 2D. At low swim pressure, the vesicle exhibits a discontinuous transition from a spherical to a prolate shape, while at high swim pressure it shows exotic spatio-temporal oscillations which we call active vesicle pearling.
In our model the active Brownian particles have a self-propulsion velocity and interact with each other and with the membrane of the vesicle via a short-ranged repulsive potential SI . The membrane is described by a Helfrich bending free energy (Helfrich, 1973), augmented by two terms that allow us to control the membrane area and the reduced volume , which is the volume of the vesicle relative to that of a sphere with the same area SI . Since lipid membranes are typically impermeable for colloidal particles and proteins of 10-100 nm in diameter, we keep the number of particles within the vesicle fixed.
Fig. 1a shows that active forces can drastically affect the shape behavior of vesicles. The figure shows the phase diagram as a function of the self-propulsion speed and the reduced volume , which can be controlled experimentally via the osmotic pressure Fettiplace and Haydon (1980). When the self-propulsion speed is zero, the system reproduces the phase behavior of vesicles in thermodynamic equilibrium Seifert (1997): as the reduced volume is decreased, the vesicle exhibits a series of shape transformations, from spherical to prolate, oblate and eventually to a stomatocyte. However, our results show that when the system is driven out of thermodynamic equilibrium, by increasing , the stomatocyte and the oblate shape loose their stability, while the prolate shape is favoured. This is due to a positive feedback between the accumulation of particles in curved regions of the membrane and the higher pressure that these particles generate, which increases the curvature. This mechanism drives the vesicle to a prolate shape in which the particles accumulate at the poles of the vesicle.
Clearly, active forces can quantitatively change the shape behavior of vesicles when their volume is controlled. However, in many experimental systems the osmotic pressure is zero, which means that the volume is free to change Fettiplace and Haydon (1980). The lines connecting the blacks dots in Fig. 1a show that in this scenario a new, discontinuous phase transition arises. These lines show the “equation-of-state”, the relation between the reduced volume and the propulsion velocity . As expected, decreases as is increased. Particles with higher propulsion velocity generate a higher force at the two poles of the vesicle, stretching the vesicle and reducing its volume. Surprisingly, however, the equation-of-state splits into two branches via a discontinuous transition. One corresponds to a shape that is still fairly spherical, while the other corresponds to a distinct prolate shape. To determine the “coexistence” velocity where both shapes are equally likely, we computed as described in (SI, ) the stationary probability distribution of the reduced volume using Forward Flux Sampling (FFS), which is a numerical technique to efficiently simulate rare events in both equilibrium and non-equilibrium systems (Allen et al., 2005; Valeriani et al., 2007). Fig. 1(b) shows the generalized free energy as a function of for three different values of . At , the prolate and spherical shapes are equally stable, yet separated by a barrier, explaining the discontinuous nature of the transition between them. At and , the prolate and spherical shapes are about to loose their stability, respectively.
This discontinuous transition between a symmetrical (circular) and non-symmetrical (elliptical) shape has not been observed in 2D systems in which active particles are confined within a semi-flexible ring polymer Tian et al. (2015); Paoluzzi et al. (2016). Since we expect that the nature of the transition depends on the competition between the elastic and the swim pressure, we analyzed these for the systems in 2D and 3D. Both in 3D (Fig. 2a) and 2D (Fig. 2b) the elastic pressure increases linearly with the mean curvature SI , suggesting that the different behaviour in 2D and 3D is due to the swim pressure.
To elucidate the origin of the scaling behavior of the swim pressure with membrane curvature, we turn to a minimal model system consisting of ideal active particles that do not interact with each other, but do interact with a curved yet static surface. The active particles are confined between two sinusoidal surfaces described by and , respectively, where is chosen to be large enough such that the correlation between the particle distributions at the two respective surfaces vanishes. The local swim pressure per unit density of the active particles is computed by simulations as described in SI .
The pressure reaches its highest value at the outer apices where the particles accumulate, and its lowest at the inner apices. Fig. 2a shows the pressure difference as a function of the absolute local mean curvature at the apices. For comparison, we also compute in a 2D system with as in Nikola et al. (Nikola et al., 2016), see Fig. 2b. Surprisingly, while grows (sub)-linearly with curvature in 2D, it grows super-linearly in 3D (Fig. 2). The latter observation is consistent with the finding of Fily et al. that the density of particles confined to the surface of an ellipsoid is proportional to the local Gaussian curvature Fily et al. (2016); Fil . The reason why the swim pressure increases superlinearly in 3D and (sub)linearly in 2D is that in 3D there is one more dimension in which the membrane is curved, such that in 3D the particles accumulate more strongly with curvature. This mechanism does indeed not rely on excluded-volume interactions. In fact, given the low particle volume fraction of about 5% at the onset of the sphere-prolate transition, they do not significantly change the scaling behavior (Fig. S1 SI ).
The different scaling of the swim pressure with curvature in 2D and 3D is key to understanding why 3D systems exhibit a discontinuous transition from a symmetrical to a non-symmetrical shape, while 2D systems do not. Because in 3D the swim pressure varies super-linearly with mean curvature while the elastic pressure scales linearly with , there exists a range of values in which the swim pressure is smaller than the elastic pressure when the curvature is below a critical value and larger than the elastic pressure when (see Fig. 2b). Hence, when the local mean curvature of a vesicle shape fluctuation is smaller than , the fluctuation tends to decay, while when it tends to grow. In this range of values, a shape fluctuation thus needs to exceed a critical size before it will spontaneously grow further. This explains why there is barrier for the formation of a curved region of macroscopic size and why the transition from the spherical to the prolate shape is discontinuous. In contrast, in 2D, the near linear dependence of the swim pressure on the curvature means that, depending on the value of , the swim pressure is either larger than the elastic pressure for (almost) all curvature values, or smaller. It is the reason why in 2D the transition from the circular to the elliptical state is continuous.
The above analysis indicates that the barrier arises from the interplay between the dependence of the elastic pressure and the swim pressure on the curvature, respectively. Since the swim pressure depends on the number of particles and the translational diffusion constant , we hypothesized that these parameters will affect the height of the barrier. Fig. 3 shows that when is increased, keeping the area of the vesicle constant, the coexistence propulsion velocity at which the prolate and spherical shapes are equally likely decreases, while the height of the barrier increases. Increasing tends to increase the total force that the active particles exert on the membrane. To compensate for this and remain at coexistence, the active force per particle—the propulsion force —must decrease, which is indeed accomplished by lowering : (see SI ). Since the force per particle is lower for lower , more particles must participate in generating the force to nucleate the shape transformation. This makes the transition more collective and increases the height of the barrier. Fig. S1 shows that the height of the barrier also increases with the translational diffusion constant , which can be explained using similar arguments (see SI ).
When the propulsion velocity is increased sufficiently beyond the sphere-prolate coexistence region (Fig. 1a), the shape dynamics become even richer. More than two clusters are typically formed, with the average number of clusters increasing with (Fig. 4a). Moreover, these clusters are highly dynamic: Fig. 4b shows that they fluctuate in number, while Supplementary Movies S1 and S2 SI shows how they emerge, vanish, and move along the vesicle, splitting into smaller clusters, and merging into larger ones. We call this behavior active vesicle pearling.
The clusters form via the interplay between the vesicle shape, the swim pressure, and the interaction between the particles. Indeed, while both finite-sized and ideal particles exhibit a discontinuous sphere-prolate transition because that arises from the feedback between the swim pressure and membrane bending (Figs. 2 and S1), active pearling requires excluded-volume interactions between the particles. Increasing the velocity, active Brownian particles with excluded-volume interactions tend to form dynamic clusters even without confinement (Tailleur and Cates, 2008; Fily and Marchetti, 2012; Redner et al., 2013). Additionally, during the elongation of the vesicle not only the density of particles increases as the volume decreases, but also the confining volume is transformed into a quasi-1d tubular shape. Both of these enhance the tendency of the particles to bump into each other and jam to form clusters. Furthermore, while clusters expand the vesicle, the concomitant increase in area must, because of the area constraint, be compensated for by making the necks thinner; the thin necks, in turn, stabilize the clusters further. This dynamic mechanism is further enhanced by the fact that an elongated vesicle in 3D has an intrinsic tendency to form a dumbbell-like shape because that minimizes the Helfrich free energy (Seifert, 1997). Both the latter mechanism and the fact that the vesicle can reduce membrane area by forming a thin neck are absent in 2D, explaining why active pearling is observed in 3D and not in 2D. Clearly, while the phenomenological behavior of active vesicle pearling bears similarities to that of a Rayleigh instability Rayleigh (1892), the physical driving forces are different.
For higher and larger system size (increasing both and vesicle size), the active pearling behavior becomes more pronounced, with clusters moving in an oscillatory yet stochastic fashion from pole to pole. Color coding the different clusters reveals that new clusters tend to be nucleated from the clusters at the vesicle poles (Supplementary Movies S3 and S4 SI ). Only collectively can the particles generate enough force to deform the thin neck and create a new “pearl”. The barrier to nucleate a new pearl increases with and (see Fig. 3 and Fig. S1 SI ), explaining why increasing these parameters makes active pearling more prominent.
In conclusion, our results show how the interplay between active forces and elastic bending forces yields phase behavior that cannot be observed in equilibrium systems Voituriez et al. (2006); Galajda et al. (2007); Wan et al. (2008); Doostmohammadi et al. (2018); Mietke et al. (2019); Tailleur and Cates (2008); Redner et al. (2013); Buttinoni et al. (2013); Keber et al. (2014). Our work paves the way for understanding the interplay between active forces and vesicle shape deformations. This could be used in controlling the deformations of synthetic vesicles for, e.g., drug delivery. Our work may also contribute to our understanding of shape transformations of living cells, which are influenced by active forces as generated by, e.g., microtubule filaments. Our predictions can be tested experimentally via vesicles containing Pt-coated colloids or bacteria (Cameron et al., 2000). We expect that the mechanical torque on the particles as exerted by the membrane and the torque between particles as arising from mechanical or hydrodynamic interactions, which all tend to align particles, will make the transition more collective and hence raise the barrier; the effect of hydrodynamic interactions between the particles and the membrane is more difficult to predict, and requires future work (Burkholder and Brady, 2018; Thutupalli et al., 2018). Other fruitful extensions would be the investigation of active particles in vesicles with an active, gel-like cortex or vesicles immersed in a bath of active particles.
This work was supported by the Netherlands Organisation for Scientific Research (NWO). We thank Martin van Hecke, Peter Bolhuis and Bela Mulder for a critical reading of the manuscript.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Cavagna et al. (2017) A. Cavagna, D. Conti, C. Creato, L. Del Castello, I. Giardina, T. S. Grigera, S. Melillo, L. Parisi, and M. Viale, Nat. Phys. 13 , 914 (2017).
- 2Kearns (2010) D. B. Kearns, Nat. Rev. Microbiol. 8 , 634 (2010).
- 3Mora et al. (2016) T. Mora, A. M. Walczak, L. Del Castello, F. Ginelli, S. Melillo, L. Parisi, M. Viale, A. Cavagna, and I. Giardina, Nat. Phys. 12 , 1153 (2016).
- 4Howse et al. (2007) J. R. Howse, R. A. L. Jones, A. J. Ryan, T. Gough, R. Vafabakhsh, and R. Golestanian, Phys. Rev. Lett. 99 , 048102 (2007).
- 5Jiang et al. (2010) H.-R. Jiang, N. Yoshinaga, and M. Sano, Phys. Rev. Lett. 105 , 268302 (2010).
- 6Wilson et al. (2012) D. A. Wilson, R. J. M. Nolte, and J. C. M. van Hest, Nat. Chem. 4 , 268 (2012).
- 7Palacci et al. (2013) J. Palacci, S. Sacanna, A. P. Steinberg, D. J. Pine, and P. M. Chaikin, Science 339 , 936 (2013).
- 8Bricard et al. (2013) A. Bricard, J. B. Caussin, N. Desreumaux, O. Dauchot, and D. Bartolo, Nature 503 , 95 (2013).