Spatial interactions and oscillatory tragedies of the commons
Yu-Hui Lin, Joshua S. Weitz

TL;DR
This paper explores how spatial interactions and feedback mechanisms can cause oscillatory tragedies of the commons, revealing new emergent dynamics and potential ways to prevent resource depletion.
Contribution
It introduces a novel framework combining stochastic and mean field models to analyze long-term feedback and spatial effects in tragedies of the commons.
Findings
Oscillatory TOC-s emerge from payoff-dependent transition rules.
Spatial interactions lead to cooperative wave fronts and cluster formation.
Emergent dynamics suggest new mechanisms for TOC prevention.
Abstract
A tragedy of the commons (TOC) occurs when individuals acting in their own self-interest deplete commonly-held resources, leading to a worse outcome than had they cooperated. Over time, the depletion of resources can change incentives for subsequent actions. Here, we investigate long-term feedback between game and environment across a continuum of incentives in an individual-based framework. We identify payoff-dependent transition rules that lead to oscillatory TOC-s in stochastic simulations and the mean field limit. Further extending the stochastic model, we find that spatially explicit interactions can lead to emergent, localized dynamics, including the propagation of cooperative wave fronts and cluster formation of both social context and resources. These dynamics suggest new mechanisms underlying how TOCs arise and how they might be averted.
Click any figure to enlarge with its caption.
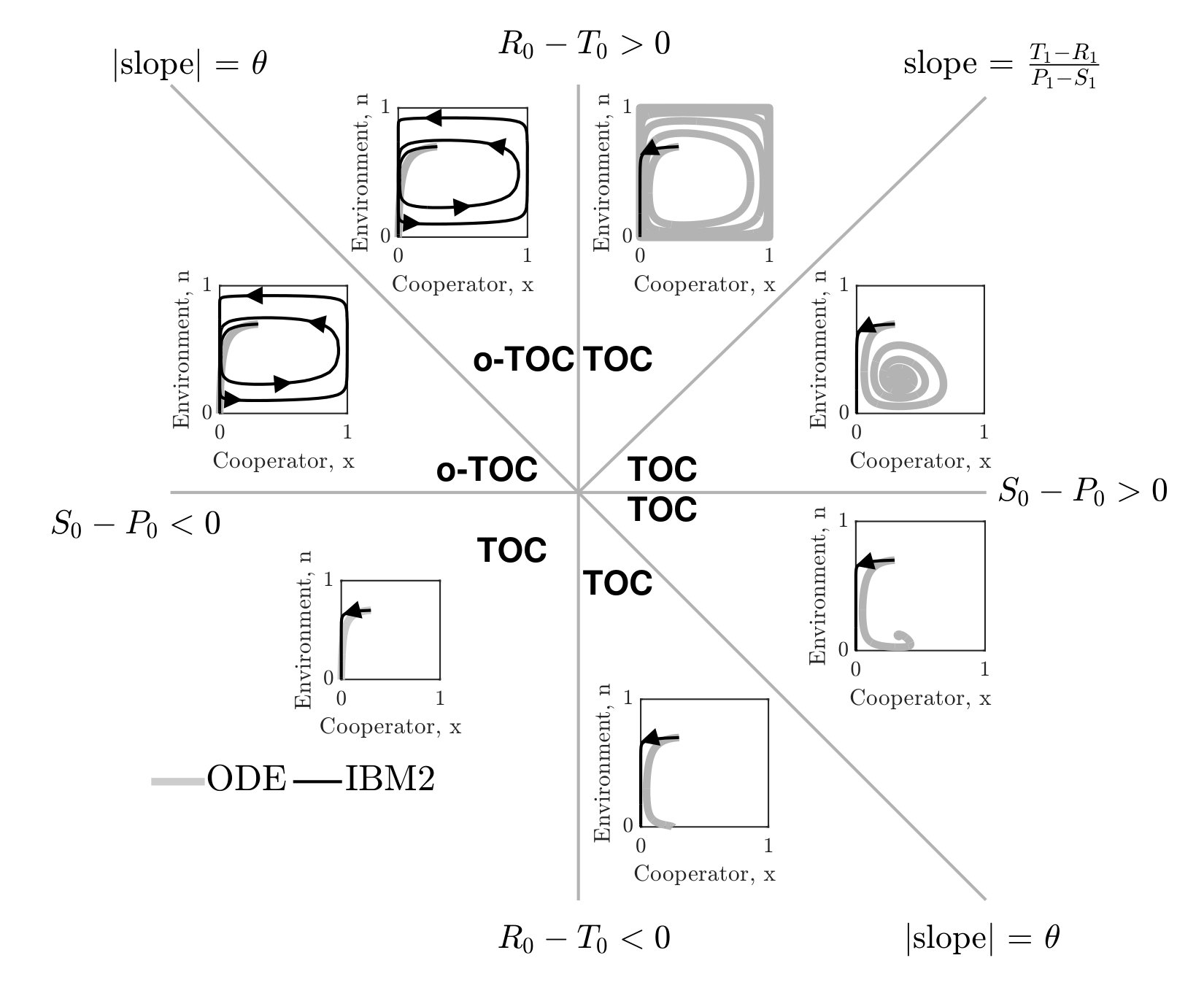 Figure 1
Figure 1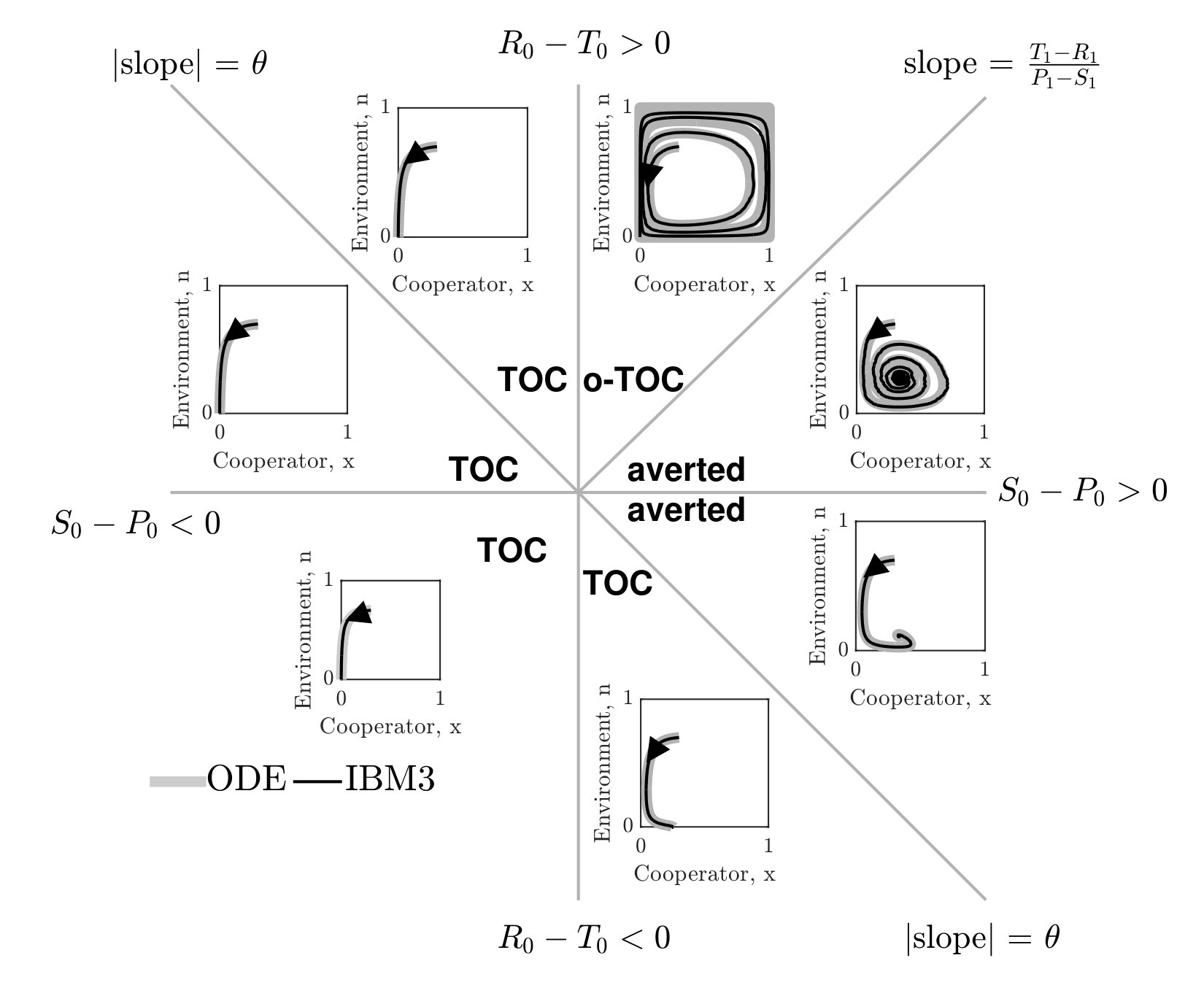 Figure 1
Figure 1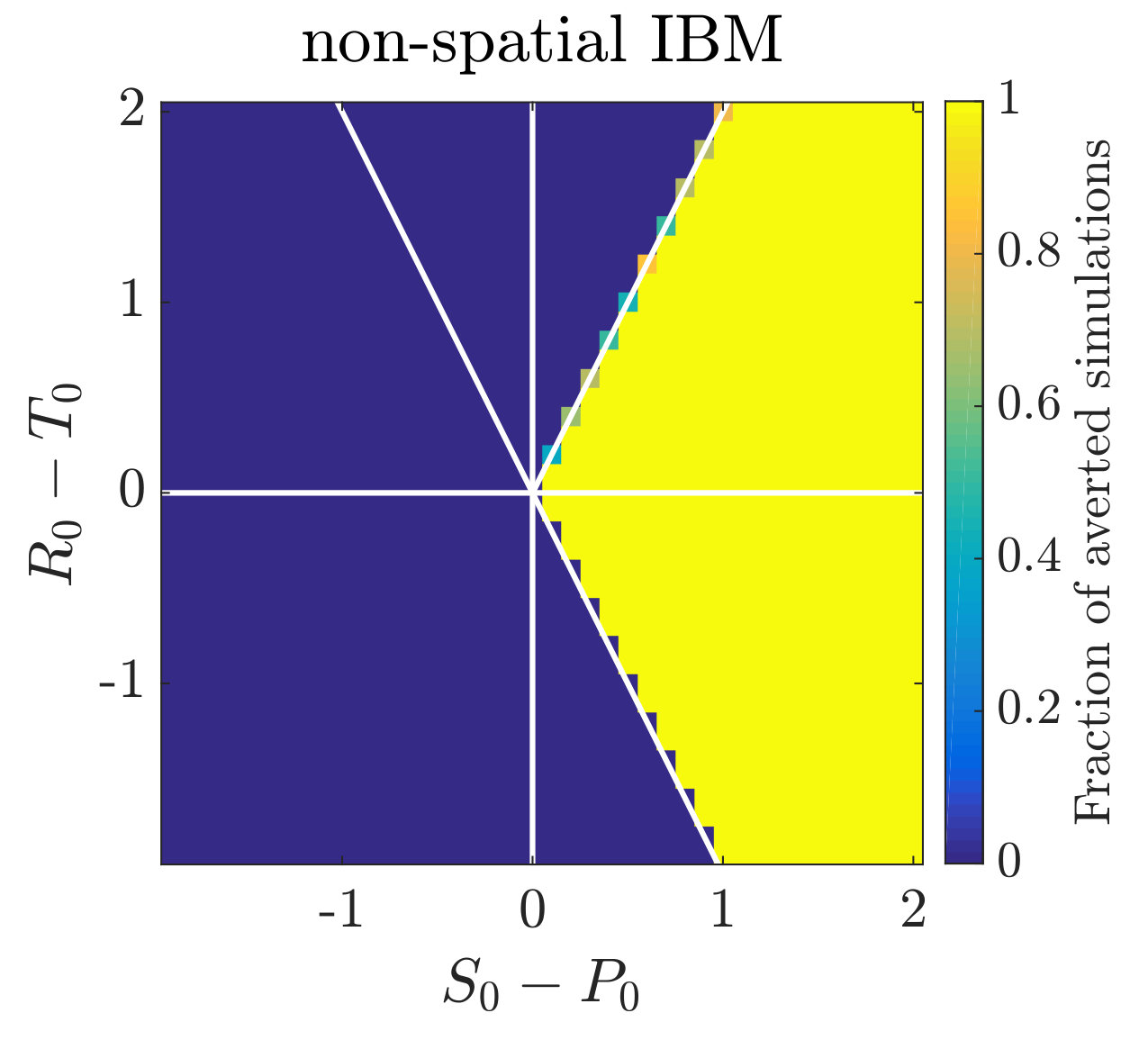 Figure 2
Figure 2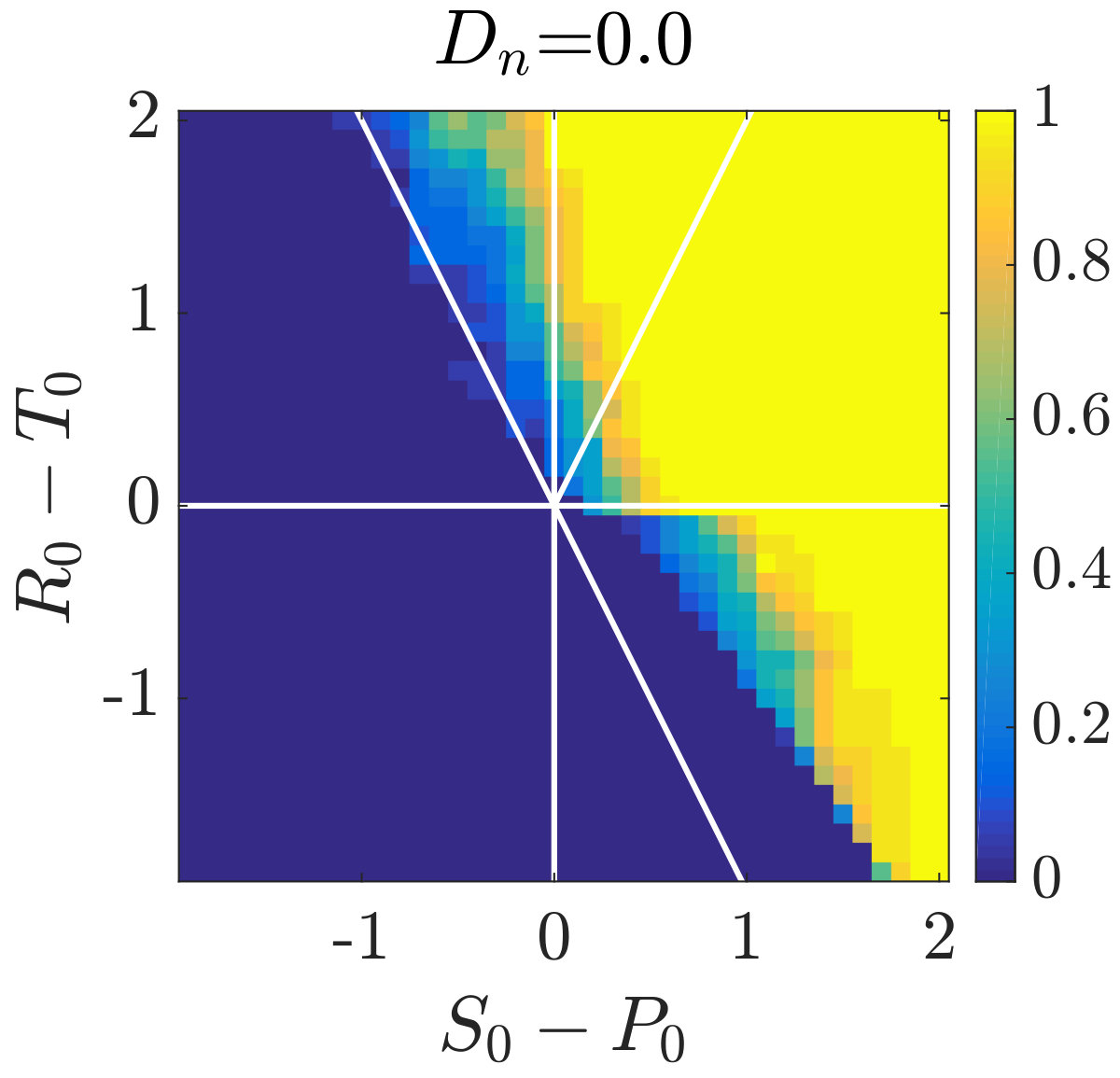 Figure 2
Figure 2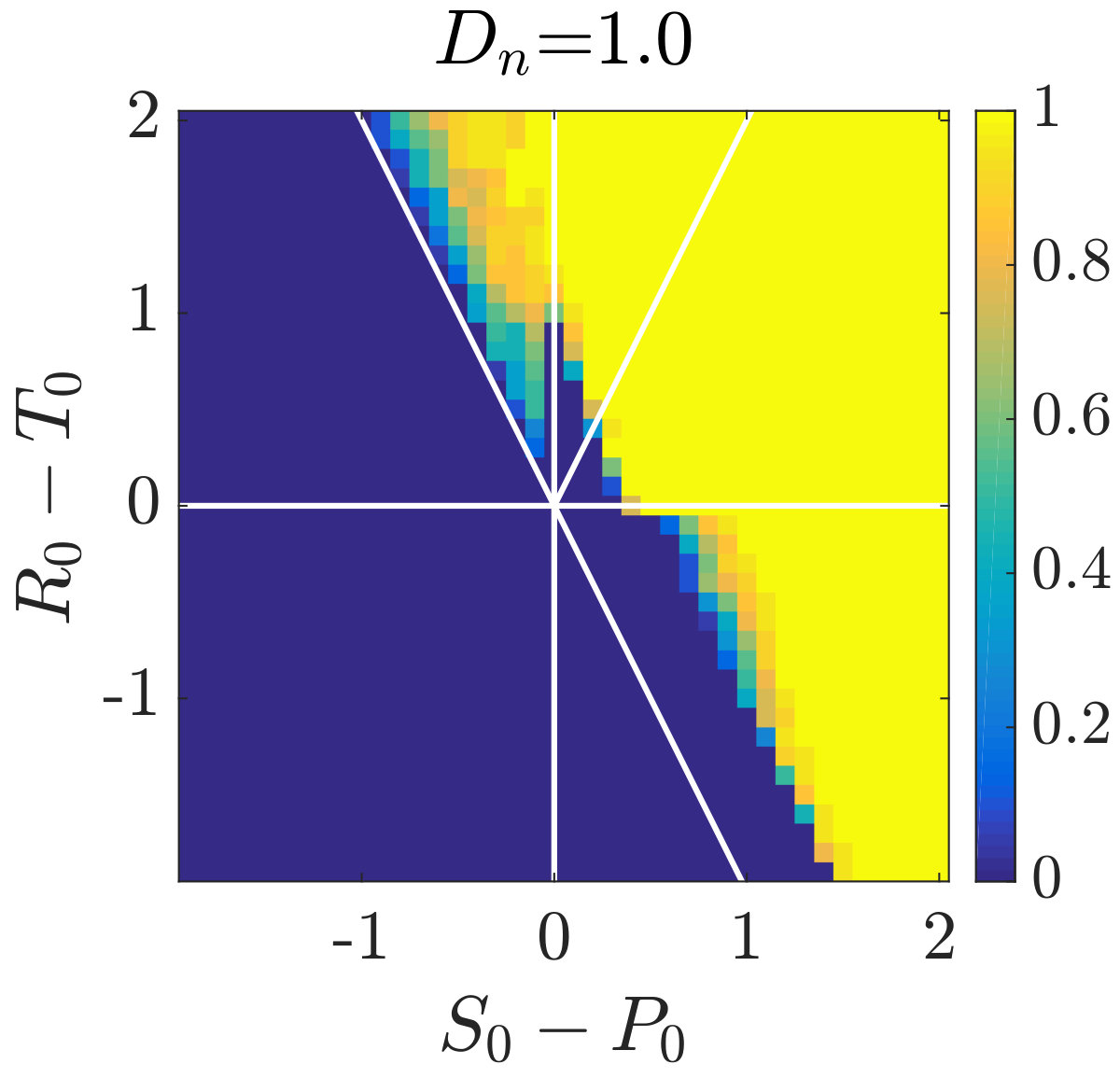 Figure 2
Figure 2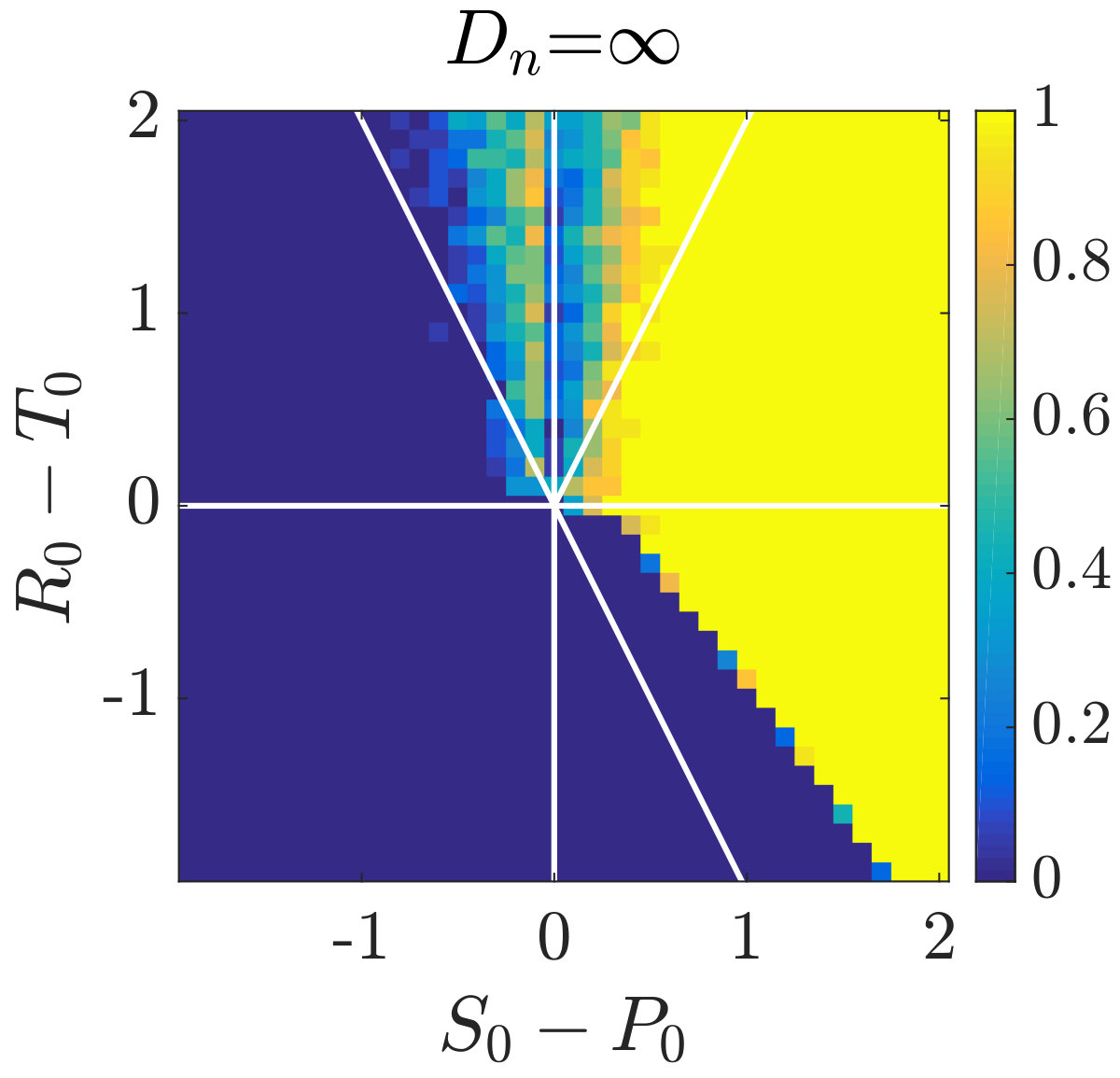 Figure 2
Figure 2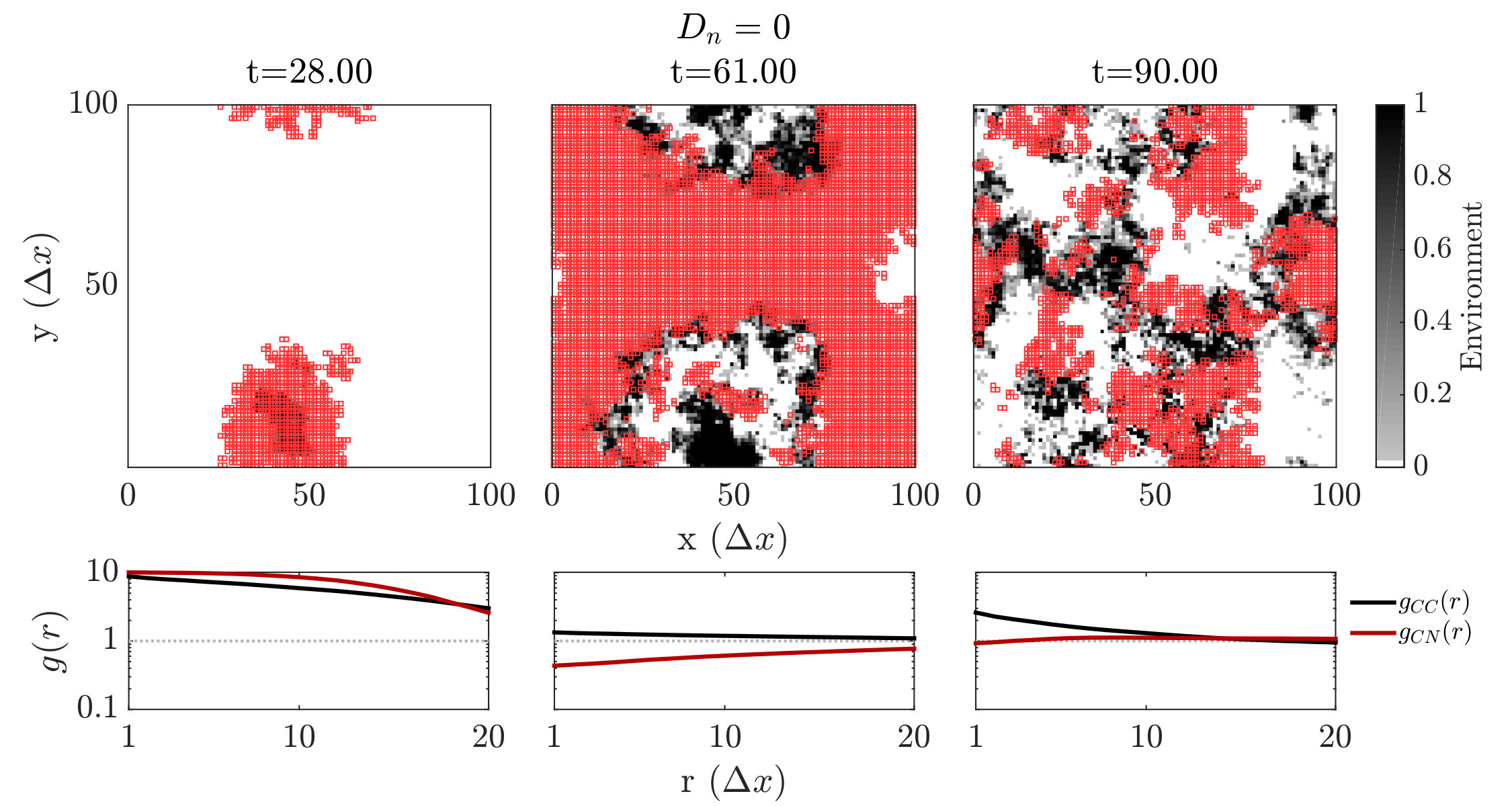 Figure 3
Figure 3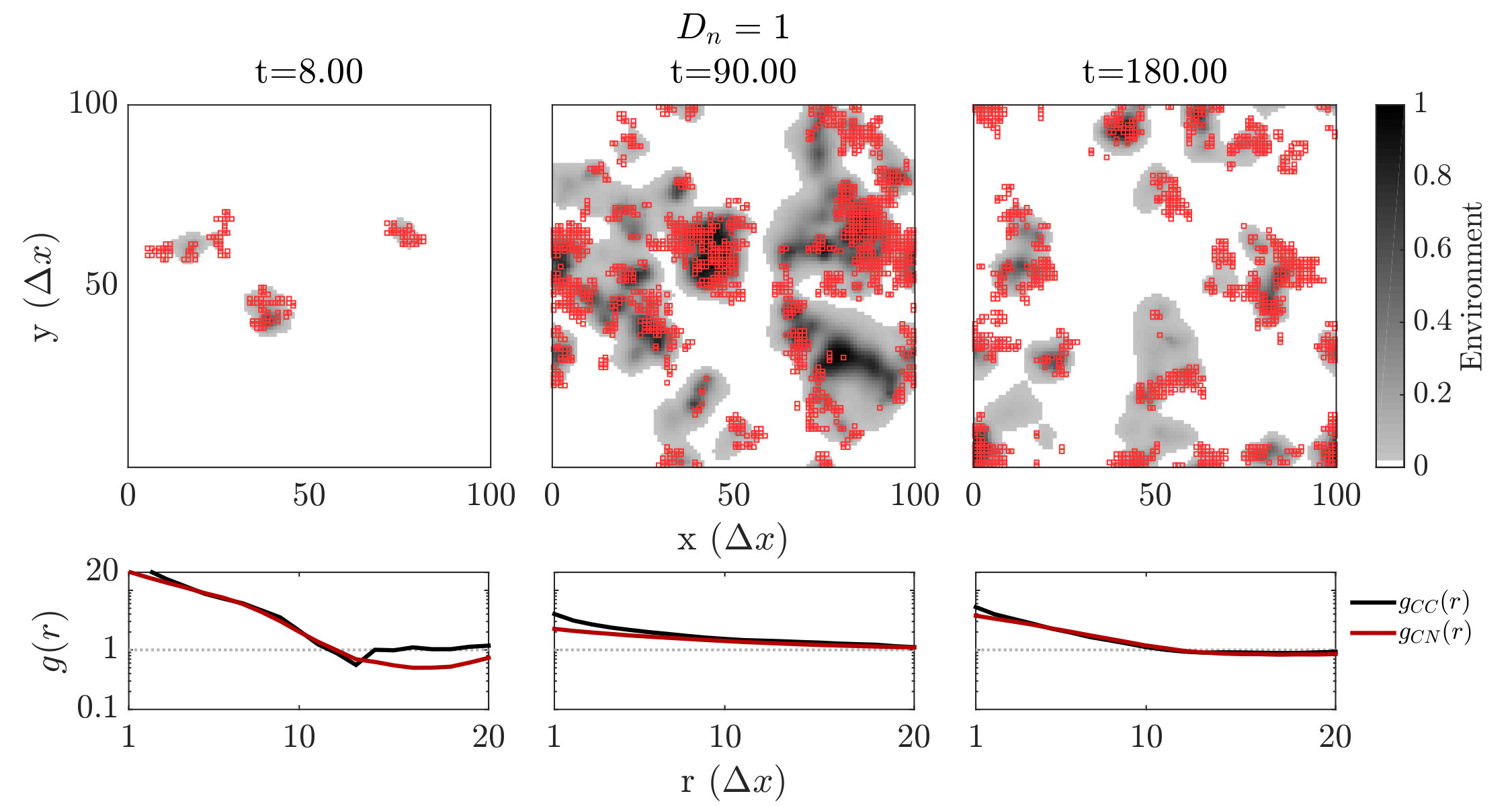 Figure 3
Figure 3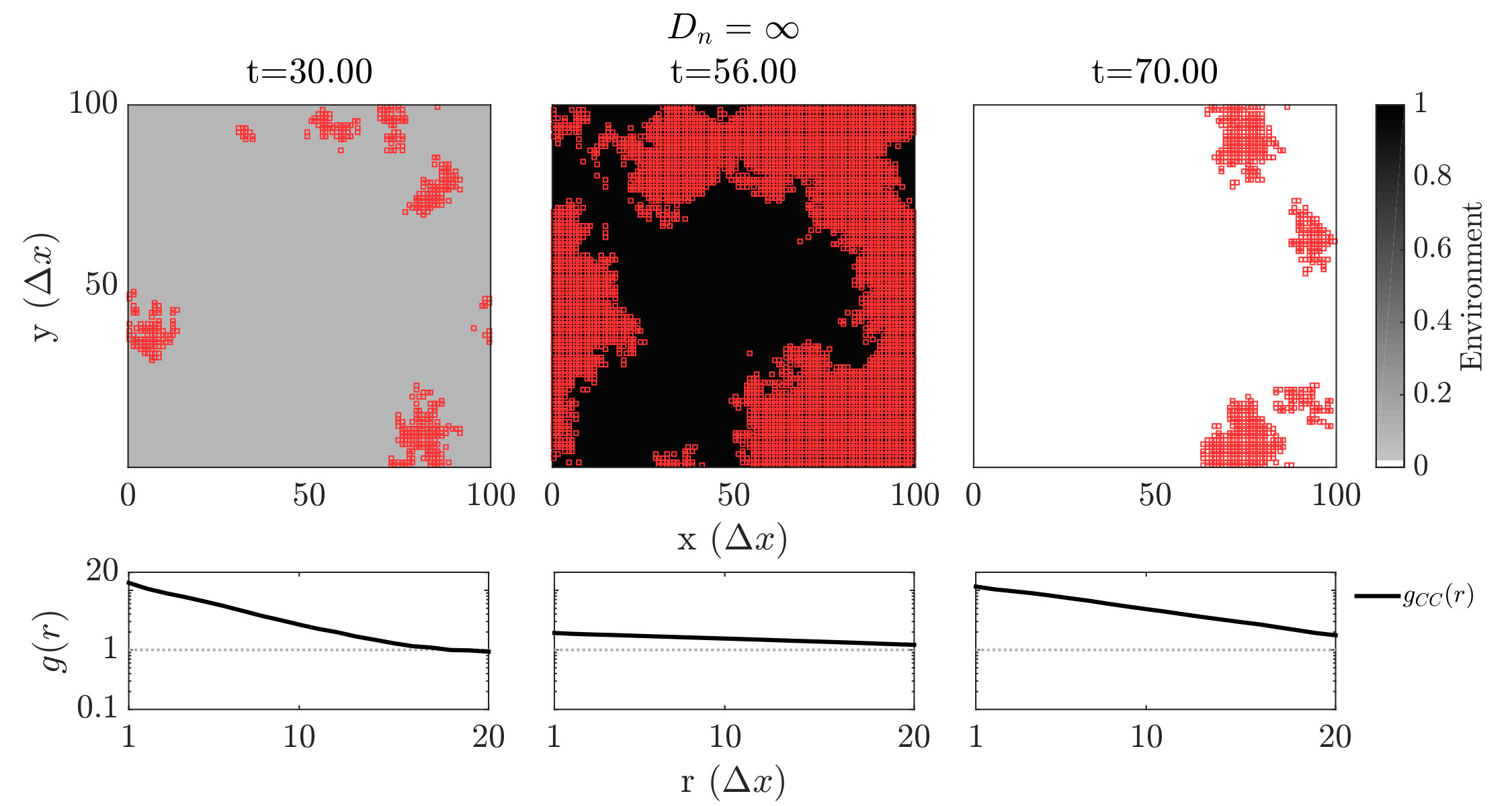 Figure 3
Figure 3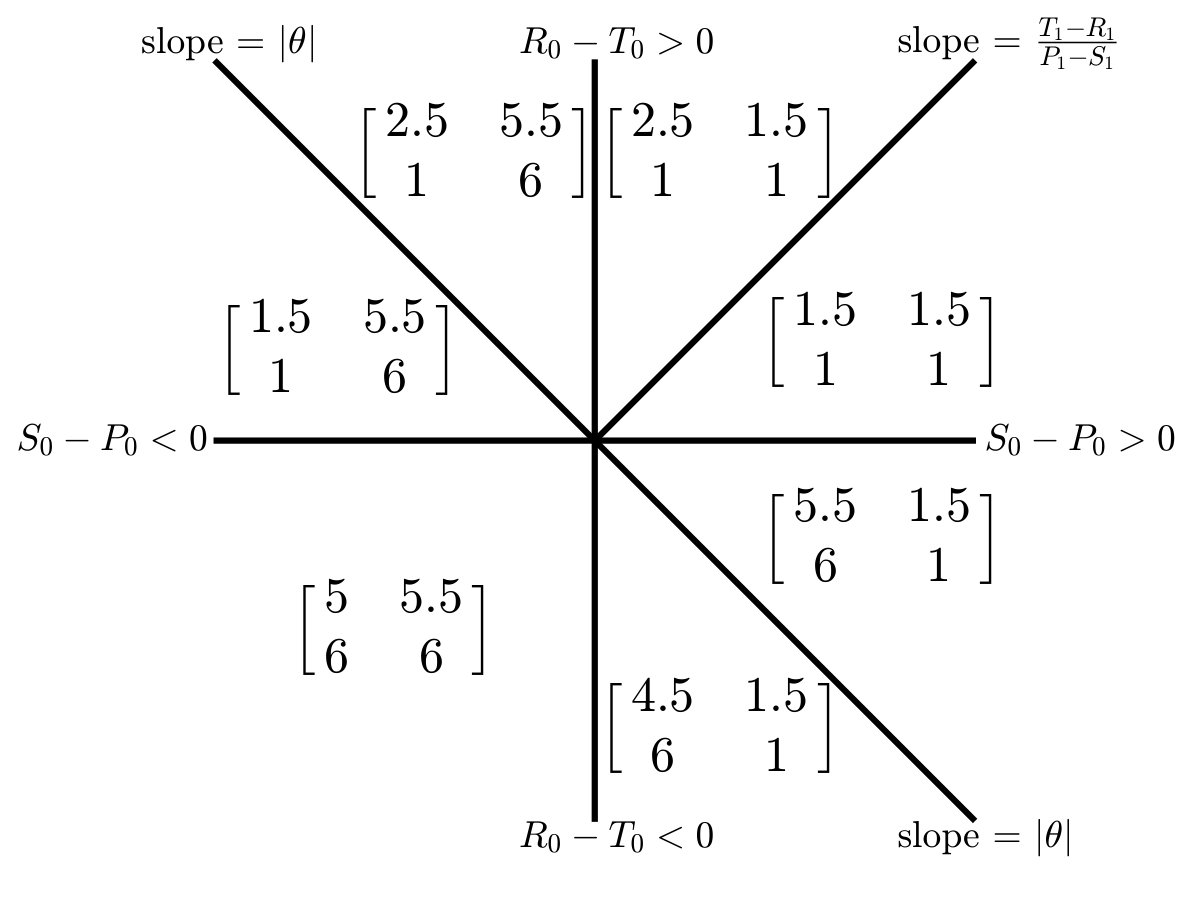 Figure 1
Figure 1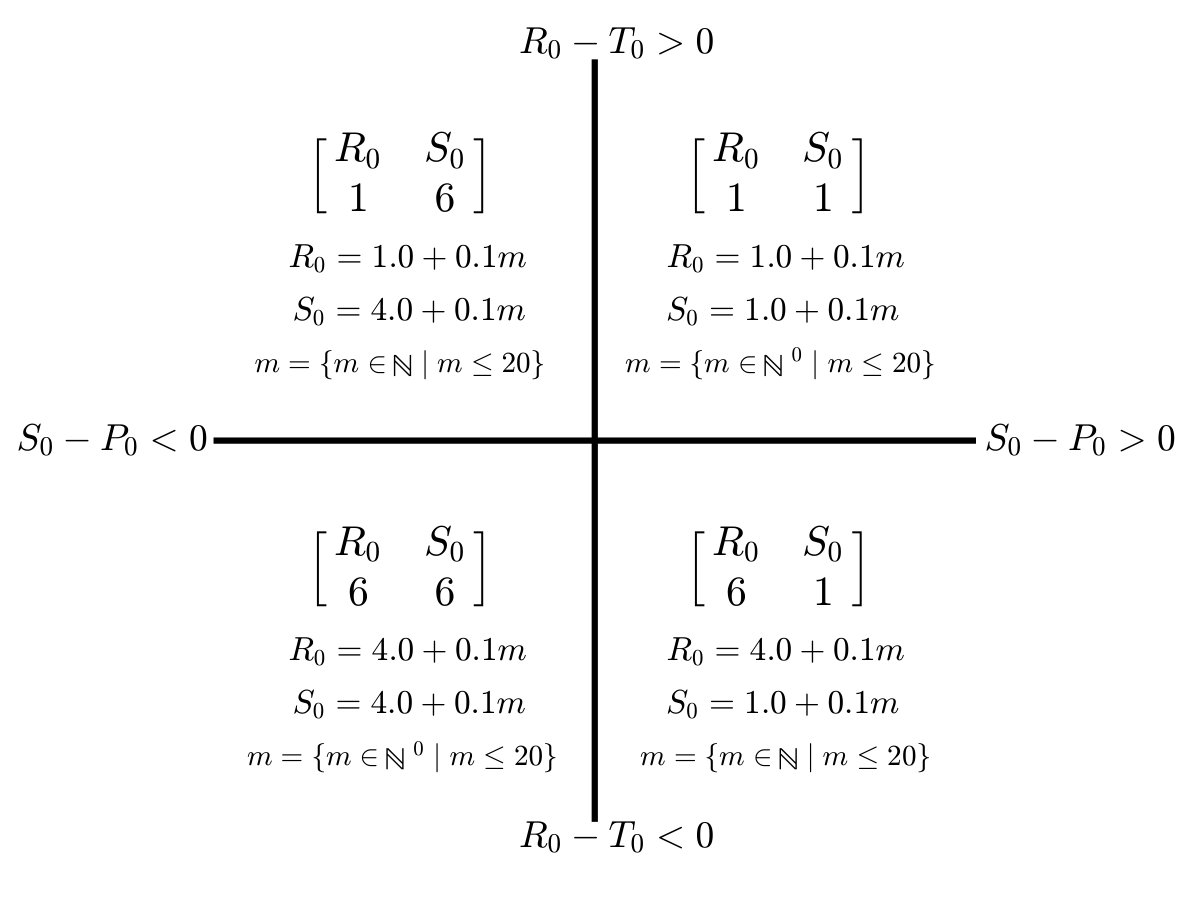 Figure 2
Figure 2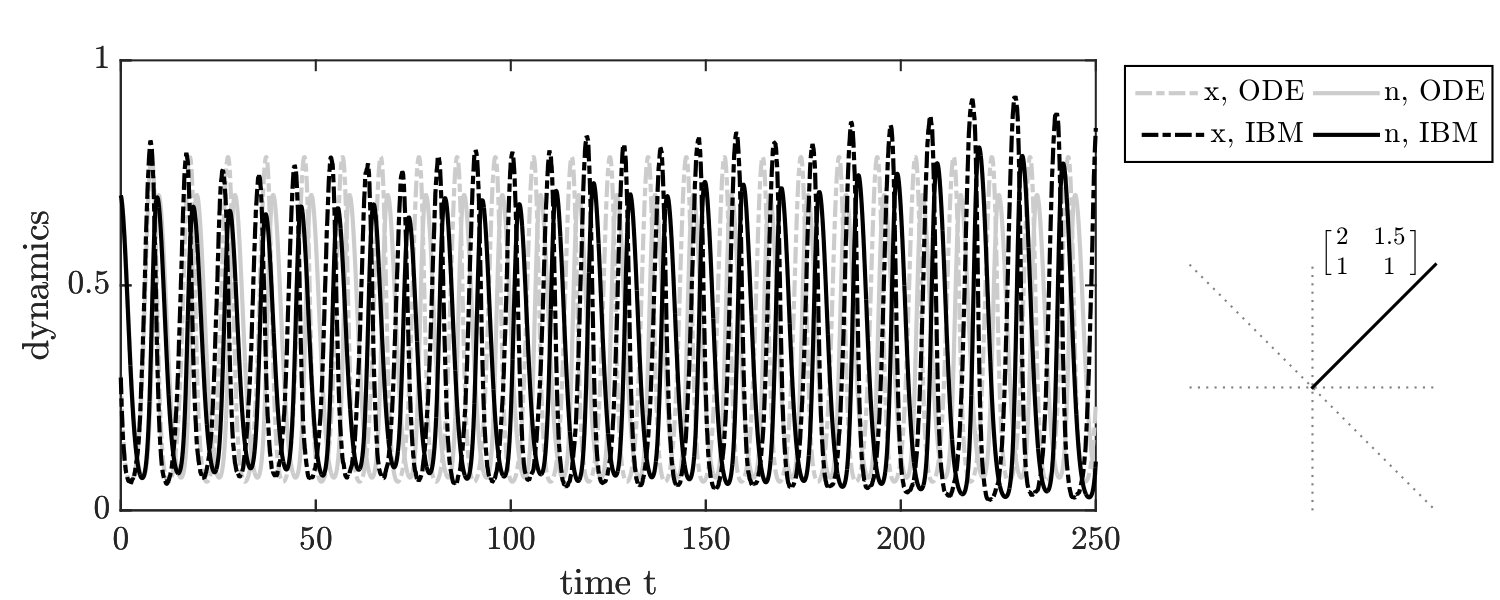 Figure 3
Figure 3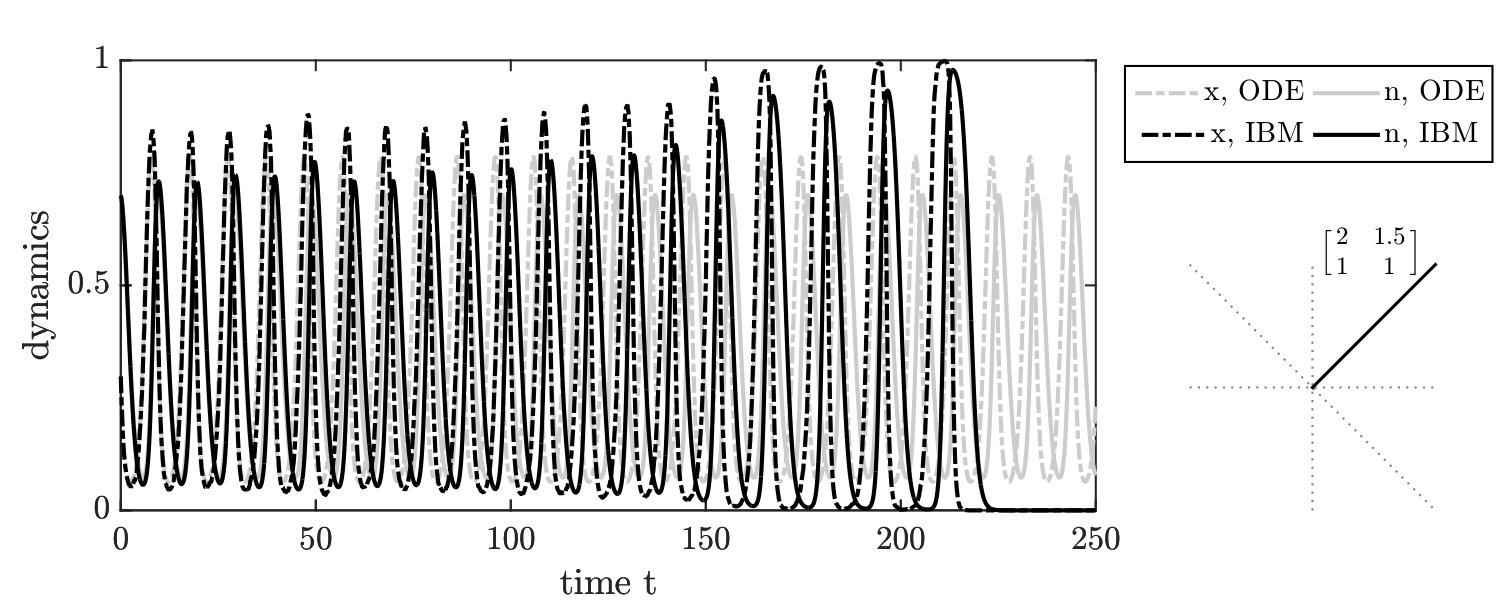 Figure 3
Figure 3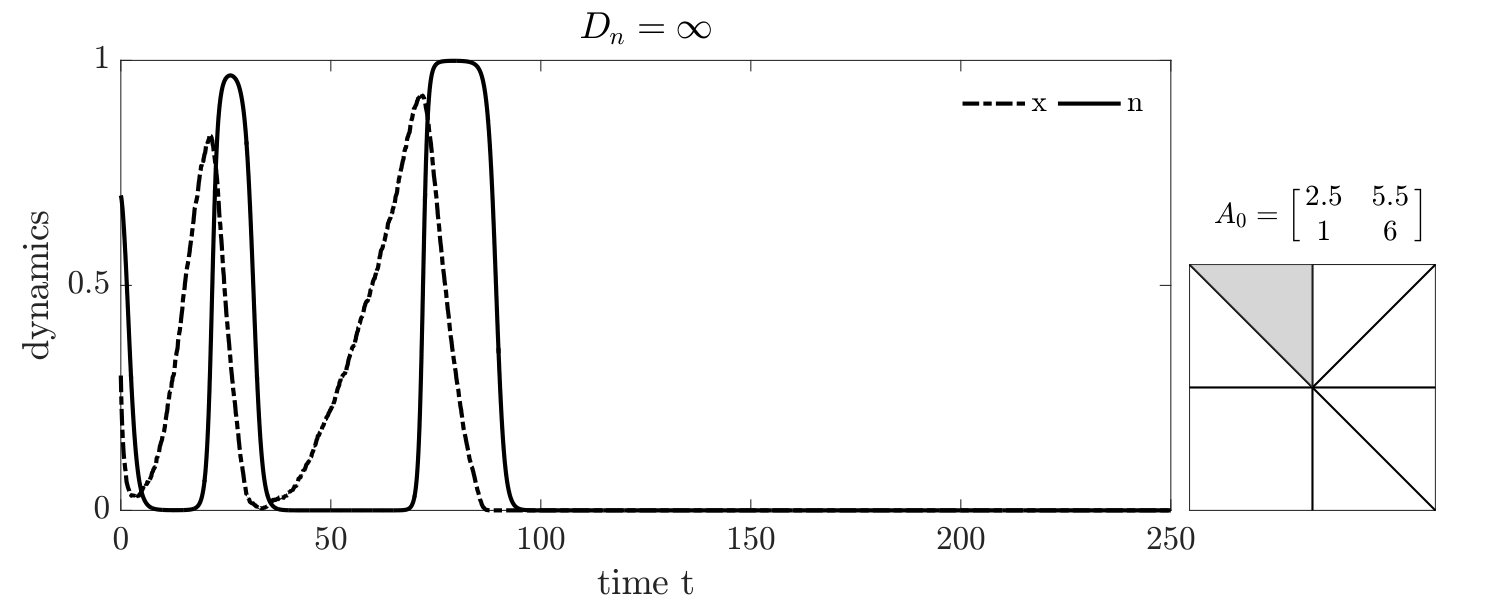 Figure 4
Figure 4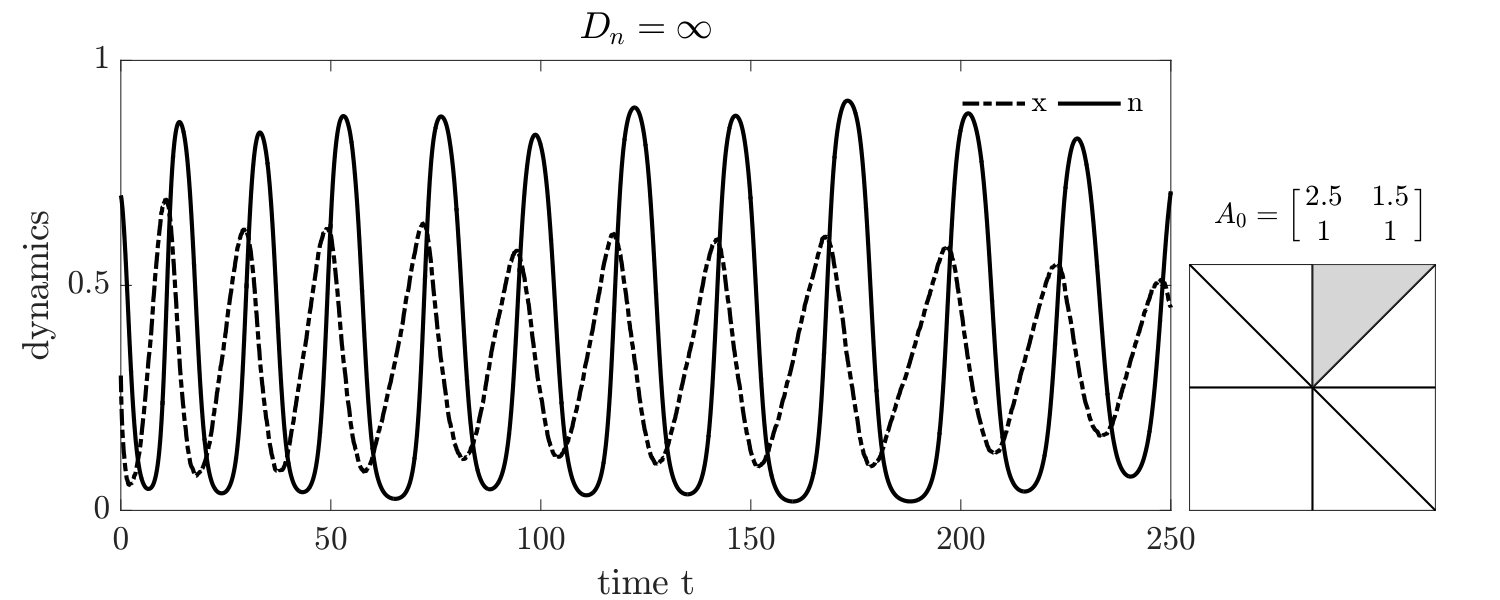 Figure 4
Figure 4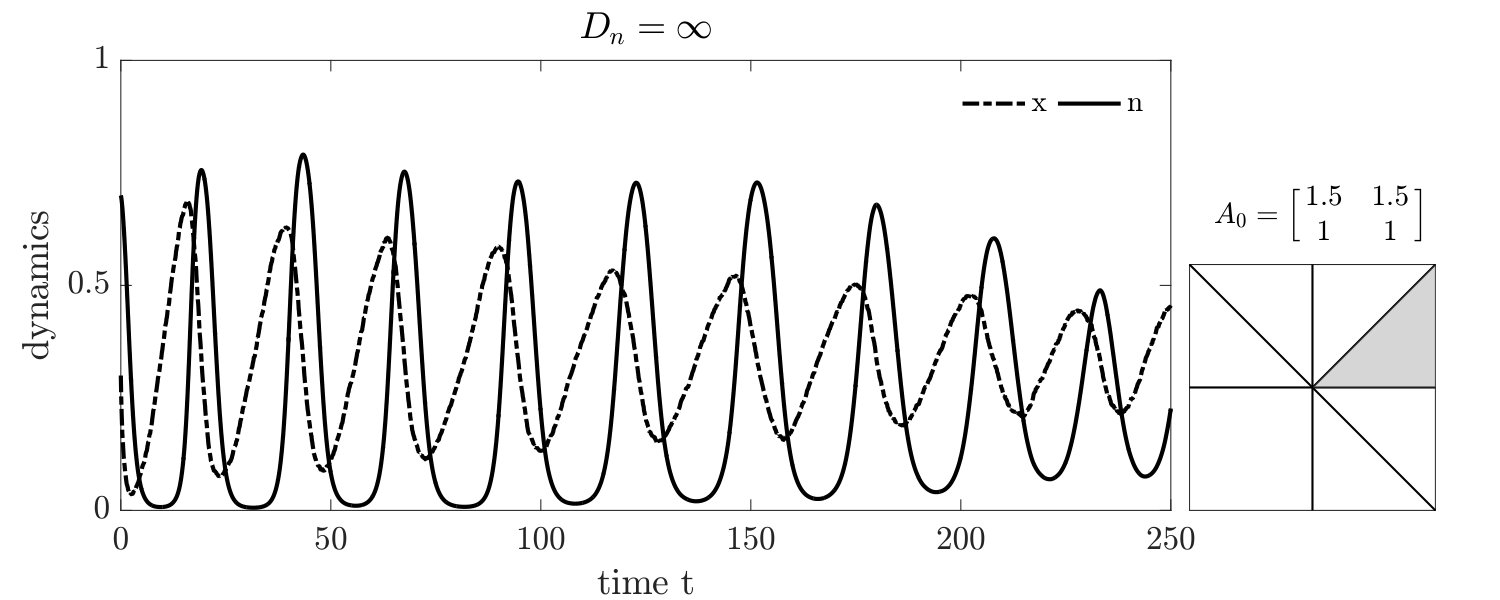 Figure 4
Figure 4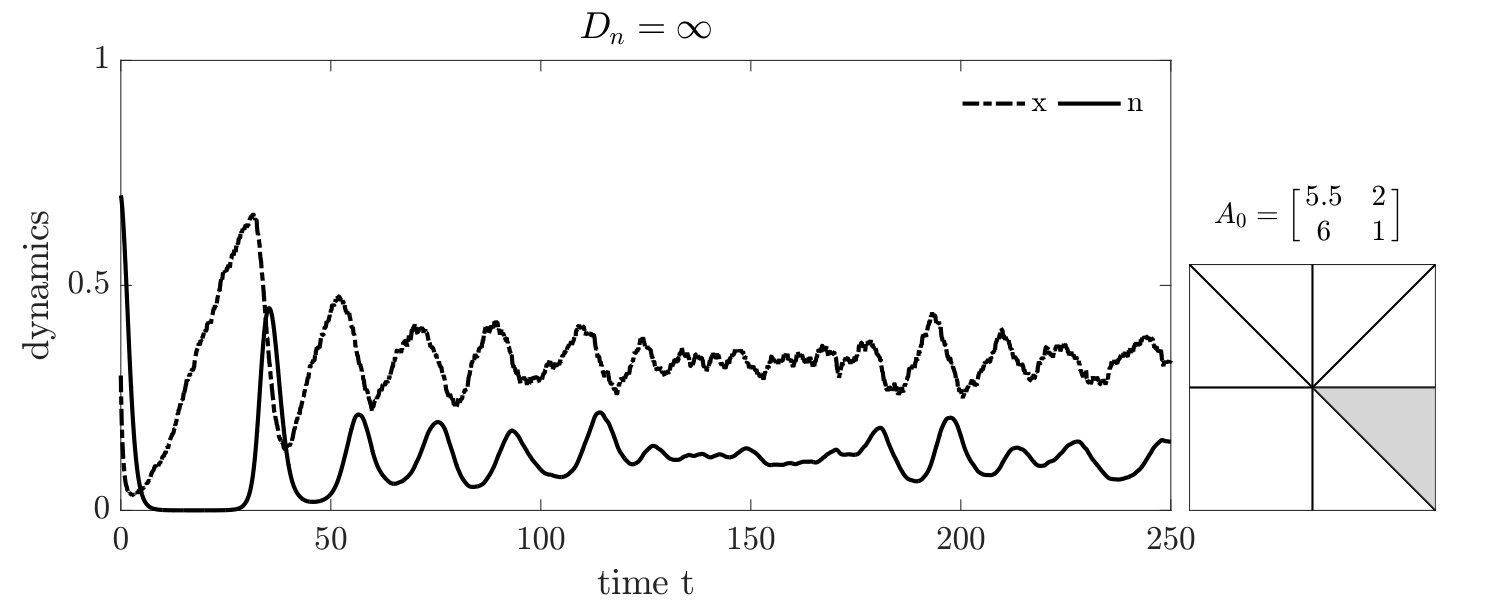 Figure 4
Figure 4Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Spatial interactions and oscillatory tragedies of the commons
Yu-Hui Lin
School of Physics, Georgia Institute of Technology, Atlanta, GA, USA
Joshua S. Weitz
School of Physics, Georgia Institute of Technology, Atlanta, GA, USA
School of Biological Sciences, Georgia Institute of Technology, Atlanta, GA, USA
Abstract
A tragedy of the commons (TOC) occurs when individuals acting in their own self-interest deplete commonly-held resources, leading to a worse outcome than had they cooperated. Over time, the depletion of resources can change incentives for subsequent actions. Here, we investigate long-term feedback between game and environment across a continuum of incentives in an individual-based framework. We identify payoff-dependent transition rules that lead to oscillatory TOC-s in stochastic simulations and the mean field limit. Further extending the stochastic model, we find that spatially explicit interactions can lead to emergent, localized dynamics, including the propagation of cooperative wave fronts and cluster formation of both social context and resources. These dynamics suggest new mechanisms underlying how TOCs arise and how they might be averted.
In 1968, Garrett Hardin explored a social dilemma, which he termed the ‘Tragedy of the Commons’ (TOC) Hardin (1968). The social dilemma arises when two individuals choose amongst distinct strategies to utilize a limited public good. Both individuals receive the maximal combined benefit if they utilize the public good with restraint, i.e., if they ‘cooperate’. However, each individual receives the maximal personal benefit if they utilize the public good without restraint, i.e., if they ‘defect’, while their opponent cooperates. As a consequence, individuals acting rationally will cheat leaving all worse off. Hardin argued that such a TOC is inevitable Hardin (1968).
Evolutionary dynamics arising from a TOC dilemma can be modeled in terms of changes in the frequencies of individuals from two populations. Indivdiuals interact and receive payoffs that depend on their strategy and the strategy of their opponent. In replicator dynamics Nowak (2006), the payoff represents a relative fitness which determines the growth of cooperators, with frequency , and of defectors, with frequency , i.e.,:
[TABLE]
The values and denote the context-dependent fitness payoff to cooperators and defectors respectively, given the payoff matrix Smith and Price (1973), , where , and where denotes the reward to cooperation, denotes the temptation to cheat, denotes the sucker’s payoff, and denotes the punishment given mutual defection. A TOC occurs when , , and . However, in contrast to standard game theory assumptions, payoffs are unlikely to remain fixed after repeated decisions that degraded commonly-held resources.
To address this issue, a recent model Weitz et al. (2016) considered dynamics arising given resource-dependent payoff matrices , which interpolate between and , the payoff matrices given deplete and replete resource states, respectively, i.e., . This model of coevolutionary game dynamics included feedback with the environmental state denoted by , such that
[TABLE]
where is a speed parameter and denotes the strength of cooperators in restoring the environment. In this coevolutionary model, the payoff matrices and can have markedly different Nash equilibria Nash Jr (1950). For example, when defection is uniformly favored when and cooperation is favored when , then the the system can exhibit a novel phenomenon termed an ‘oscillatory tragedy of the commons’ (o-TOC). An o-TOC denotes a trajectory in the phase plan that approaches a heteroclinic cycle. Given a replete environment, the population rapidly switches from cooperation to defection, which then degrades the environment. In the depleted environment, cooperators re-establish, improving the environment, then defectors invade and the cycle repeats. Other outcomes, including a TOC and the aversion of a TOC can emerge given other payoff matrices Weitz et al. (2016).
This coevolutionary game model is the basis for our development and analysis of an individual-based framework to assess the influence of noise (first) and spatially explicit interactions (second) on the emergent dynamics of social context and resources. To begin, consider a system comprised of cooperators and defectors, such that . A single time step consists of events. In each event, a randomly chosen individual (the focal player) interacts with another individual (the opponent) chosen at random. The payoff to the focal player influences its probability to reproduce. Critically in our proposed framework, successful reproduction by the focal player replaces a randomly chosen third individual (see Bauer and Frey (2018) for a related public goods model that decouples interaction and reproduction). The following reactions denote those transitions that lead to a change in the number of cooperators or defectors:
[TABLE]
where denote reproduction rates.
In the three-individual framework, the master equation for the dynamics of cooperators is:
[TABLE]
where the transition rates are:
[TABLE]
In the SI, we derive the expected mean field dynamics for the frequency of cooperators from the master equation:
[TABLE]
We recover the replicator dynamics of the coevolutionary model when , , , and . Hence, transition rates are a function of resource- and social-context dependent payoffs. In contrast, mean field dynamics derived via a two-player individual based model formulation (IBM2) result in a logistic dependency on distinct from the cubic nonlinearity in Eq. 7 (see SI for derivation and details).
In order to further evaluate stochastic dynamics of the IBM formulation, we simulated the joint dynamics of resources and social context using individuals. A single time step over an interval includes game steps followed by changes in resource levels, according to Eq. 3 (see Supplementary Information (SI) for details). Given the master equation analysis, we define reproduction rates based on the current environmental state . Consistent with our finding from the master equation, the simulation results of the individual-based model involving three players (IBM3) recapitulate predictions of the mean-field replicator dynamics model (see FIG. 1-right). Specifically, we identify seven distinct phases corresponding to the relative magnitude of payoffs given the resource deplete state. The phases and their asymptotic behavior agree qualitatively with mean-field predictions. In contrast, if the focal player reproduces and replaces the opponent (as is often assumed in two-player variants of spatial games), then the individual-based simulations diverge from predictions (see FIG. 1-left) as anticipated from expected mean field dynamics (see SI).
There are two notable quantitative differences in the IBM3 simulations with respect to predictions from replicator dynamics. First, whereas mean-field dynamics predict convergence to a heteroclinic cycle (see ‘o-TOC’ region in FIG. 1-right), the IBM simulations stochastically reach an absorbing state on the boundary. Such a result is anticipated in any finite size simulation, given that heteroclinic cycles asymptotically approach the boundary. Second, the mean field model predicts closed period orbits given certain symmetric properties of and (corresponding to the line with slope in FIG. 1-right.) In contrast, the IBM simulations have demographic noise, which can lead to repeated oscillations and convergence to a boundary (see FIG. S3).
To study the combined effects of spatial structure and demographic noise (see Durrett and Levin (1994)) we extended the IBM3 framework a 2-dimensional fully occupied lattice with sites per dimension given periodic boundary conditions, where the individuals are either cooperators or defectors. The focal player is selected at random and opponent is chosen randomly from the von Neumann neighborhood of the focal player. We denote the position of the focal player (opponent) as (). The focal player reproduces with probability rate given the strategy set of focal player and opponent, and , and the average local environment, . Environmental state dynamics are augmented by diffusion, i.e.,:
[TABLE]
The diffusivity controls the redistribution of resources relative to population dynamics.
Simulations of coevolutionary game-environmental dynamics reveal dramatic changes in outcomes given spatially explicit interactions. FIG. 2 compares dynamics of non-spatial and spatial IBM models with three different diffusivities, , classifying outcomes based on whether there is a TOC or not (the latter we term averted, see SI for criteria). The heat maps show the proportion of averted cases among all replicates. Spatial interactions enable TOC aversion when cooperation is favored given a coordination game context ( and , see upper left). However, spatial interactions also restrict the parameter regimes where a TOC can be averted given an anti-coordination game context ( and , see bottom right). For long-term dynamics, we find that oscillating dynamics are typical in cases (see examples in the SI). Such oscillatory dynamics can spiral inwards when TOC-s are averted or outwards to the boundary. Of note, amongst IBM models we only observe a persistent o-TOC when ; indicating the role of strong spatial coupling to induce oscillations.
We further investigated spatiotemporal dynamics focusing on variation in given parameter regimes with both averted and TOC dynamics. These regimes correspond to the case where , and where (see bottom panels of FIG. 2). The results of spatially explicit IBM3 model simulations are shown in FIG. 3 for and . Notably, all cases appear to exhibit clustering amongst cooperators and the cases with heterogeneous environmental dynamcis ( and ) also appear to exhibit clustering between cooperators and environmental resource state. However, there are markedly different types of emergent spatial patterns give variation in the diffusivity of environmental resource state. In order to assess clustering quantitatively, we analyzed the joint structure of social context and resource levels by measuring the spatial cross-correlation function:
[TABLE]
and the spatial autocorrelation function of cooperator clustering:
[TABLE]
where , and denotes the number of lattice sites within this range in both cases. We then fit the short-range components of the observed correlation at a fixed time point to a decaying exponential, i.e., given pre-factor and correlation length .We then fit the short-range components of the observed correlation at a fixed time point to a decaying exponential, i.e., given pre-factor and correlation length . The spatial autocorrelation analysis confirms the emergence of clustering amongst cooperators when the TOC is averted, i.e., for (see black lines in the sub-panels of FIG. 3). Yet there are marked differences in the dynamics of the cross-correlation between cooperators and the environmental state.
For , the environment and cooperative population propagate outward as a wave. The cooperative population spread leaving patches of resource replete environments. The plots shows that and can be positively correlated as a wave initiates but negatively correlated once defectors invade and replace resource replete environments, leading to (often disjoint) patchy distributions of both resources and cooperators. In contrast, for , small clusters of cooperators and localized resources form after initial transient dynamics. This feature is captured by the analysis, revealing strongly elevated cross-correlation (see the middle row of FIG. 3) as well as similar pattern in the dynamics of and . We note that these ‘gangs’ of cooperators and their environmental ‘tail’ are chased by a dominant group of defectors (see (Szabó and Tőke, 1998) for related findings in evolutionary PD models withoout environmental feedback). Finally, given , the resources are uniform across space. Cooperative clusters grow towards system sizes due to the strong spatial coupling mediated via fast resource diffusivity. The single large cooperator cluster expands and shrinks over time with increasing amplitude, as evidenced by the elevated autocorrelation of in the bottom row of FIG. 3, with rapid switches in resource state, leading to an eventual collapse of the cooperator population. We do not report given the uniform distribution of resources given .
In summary, we have developed an individual-based framework to incorporate the analysis of demographic noise and spatial interactions in coevolutionary game dynamics that coupled individual strategies and the environment Durrett and Levin (1994). The IBM involving 3 players in a game recapitulates and generalizes earlier findings from a coevolutionary game model, including the emergence of an oscillatory tragedy of the commons. Spatial interactions can shift the domains in which a tragedy of the commons may arise when compared to non-spatial models Halatek and Frey (2018). Spatially explicit dynamics also lead to novel, coherent spatiotemporal patterns Nowak and May (1992); Butler and Goldenfeld (2009, 2011); Nowak and May (1993); Nanda and Durrett (2017), including diffusive clusters, flickering, and wave-like patterns. These joint dynamics of resources and social strategies suggest multiple avenues for future study, including formally deriving effective PDEs to characterize whether the system permits propagating waves in the large system limit. It will also be critical to evaluate the extent to which spatial interactions modify strategy-environment feedback in proposed generalizations Tilman et al. (2018) of the replicator framework underlying the present work Weitz et al. (2016) and in stochastic games with feedback between behavior and public good states (Hilbe et al., 2018). Finally, the spatial approach may also aid efforts to understand how microorganisms produce and utilize public goods, e.g., siderophores – extracellular iron harvesting enzymes Cordero et al. (2012); Niehus et al. (2017); Bauer and Frey (2018); Menon and Korolev (2015); Lewin-Epstein et al. (2017). Given increasing pressures on limited resources, we intend to leverage prior work on controlling mean-field strategy-environment dynamics Paarporn et al. (2018) to identify ways in which local manipulation of resources, strategies, and/or perceptions can help stabilize and conserve the commons.
Acknowledgements
We thank Erol Akçay, Marianne Bauer, Ceyhun Eksin, Guanlin Li, Keith Paarporn, William Ratcliff, and Peter Yunker for feedback and Stephen Beckett for reviewing code. This work was supported by the Simons Foundation (SCOPE award ID 329108, J.S.W.) and by Army Research Office grant W911NF-14-1-0402.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Hardin (1968) G. Hardin, Science 162 , 1243 (1968) . · doi ↗
- 2Nowak (2006) M. A. Nowak, Evolutionary Dynamics (Harvard University Press, 2006).
- 3Smith and Price (1973) J. M. Smith and G. R. Price, Nature 246 , 15 (1973).
- 4Weitz et al. (2016) J. S. Weitz, C. Eksin, K. Paarporn, S. P. Brown, and W. C. Ratcliff, Proceedings of the National Academy of Sciences 113 , E 7518 (2016) . · doi ↗
- 5Nash Jr (1950) J. F. Nash Jr, Econometrica: Journal of the Econometric Society , 155 (1950).
- 6Bauer and Frey (2018) M. Bauer and E. Frey, Physical Review E 97 , 042307 (2018).
- 7Durrett and Levin (1994) R. Durrett and S. Levin, Theoretical Population Biology 46 , 363 (1994).
- 8Szabó and Tőke (1998) G. Szabó and C. Tőke, Physical Review E 58 , 69 (1998).