Suppression of macroscopic oscillations in mixed populations of active and inactive oscillators coupled through lattice Laplacian
Ikuhiro Yamaguchi, Takuya Isomura, Hiroya Nakao, Yutaro Ogawa,, Yasuhiko Jimbo, Kiyoshi Kotani

TL;DR
This paper develops a theoretical framework to understand how mixed populations of active and inactive oscillators can have their synchronized oscillations suppressed through local diffusive coupling, with applications to epileptic seizure modeling.
Contribution
It introduces an approximate stability criterion based on the system's free energy and highlights the role of effective wavenumber and spatial arrangement in suppression.
Findings
Effective suppression depends on the spatial arrangement of oscillators.
The theory is validated with a cortico-thalamic model of epilepsy.
Suppression is influenced by population ratio and intensity.
Abstract
We consider suppression of macroscopic synchronized oscillations in mixed populations of active and inactive oscillators with local diffusive coupling, described by a lattice complex Ginzburg-Landau model with discrete Laplacian in general dimensions. Approximate expression for the stability of the non-oscillatory stationary state is derived on the basis of the generalized free energy of the system. We show that an effective wavenumber of the system determined by the spatial arrangement of the active and inactive oscillators is an decisive factor in the suppression, in addition to the ratio of active population to inactive population and relative intensity of each population. The effectiveness of the proposed theory is illustrated with a cortico-thalamic model of epileptic seizures, where active and inactive oscillators correspond to epileptic foci and healthy cerebral cortex tissue,…
Click any figure to enlarge with its caption.
 Figure 1
Figure 1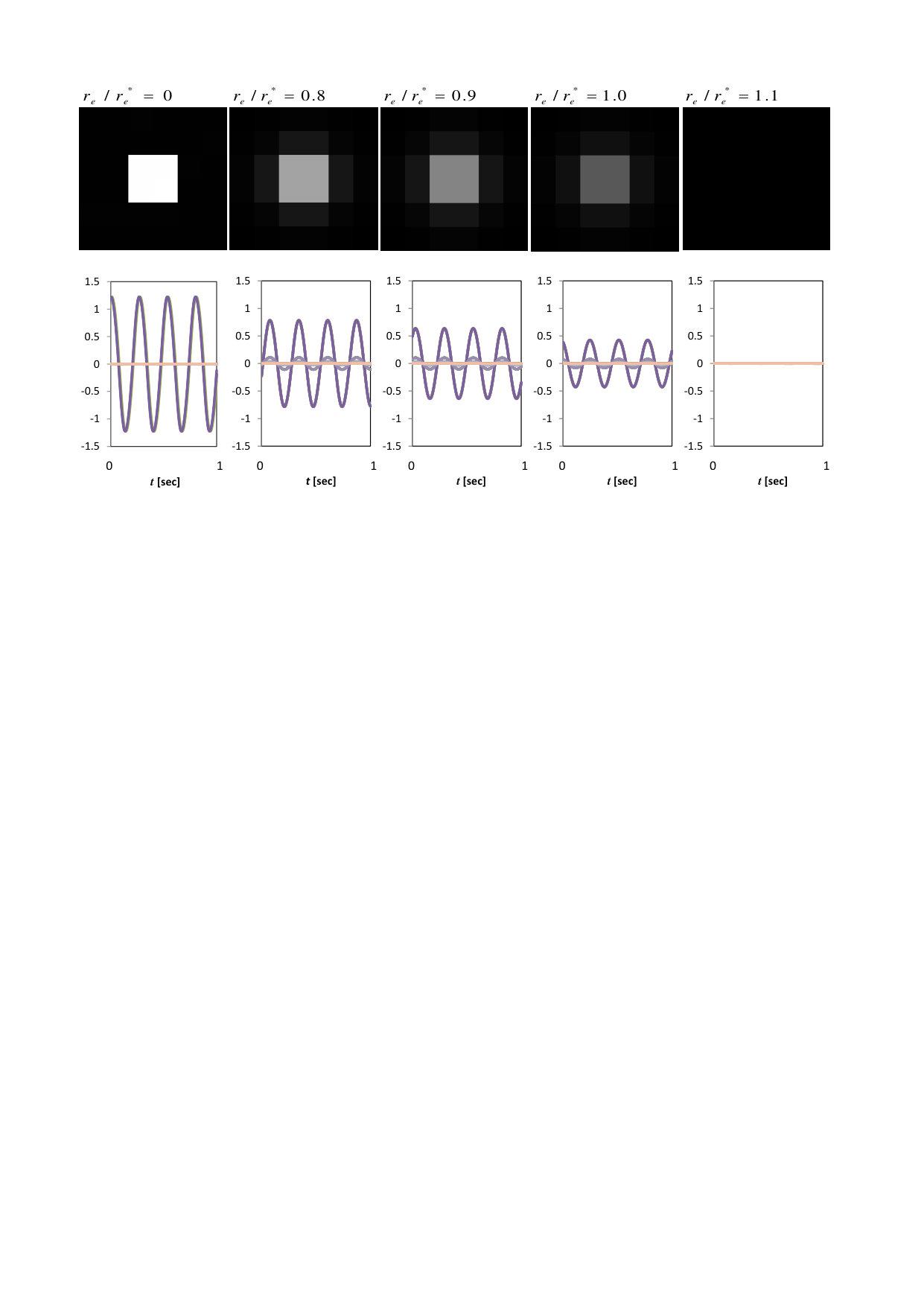 Figure 2
Figure 2 Figure 3
Figure 3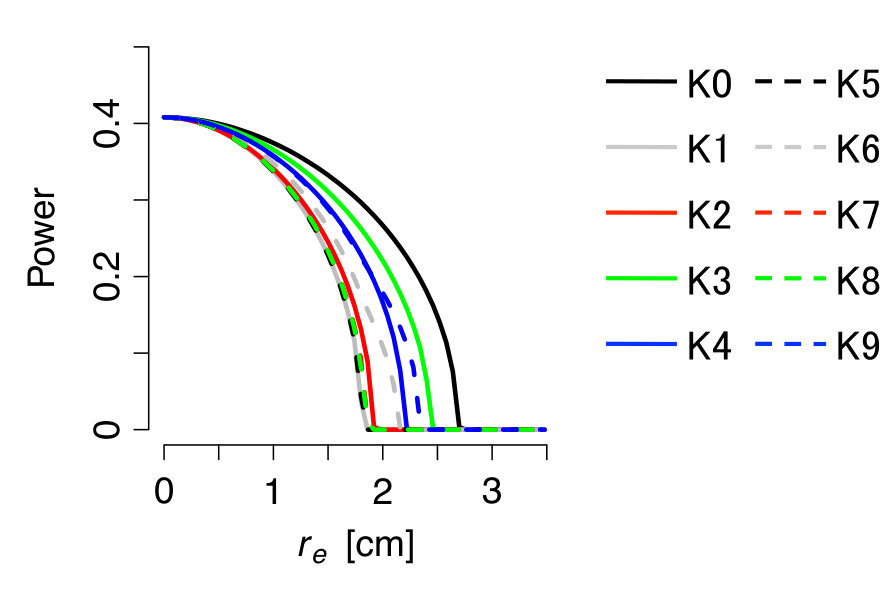 Figure 4
Figure 4 Figure 5
Figure 5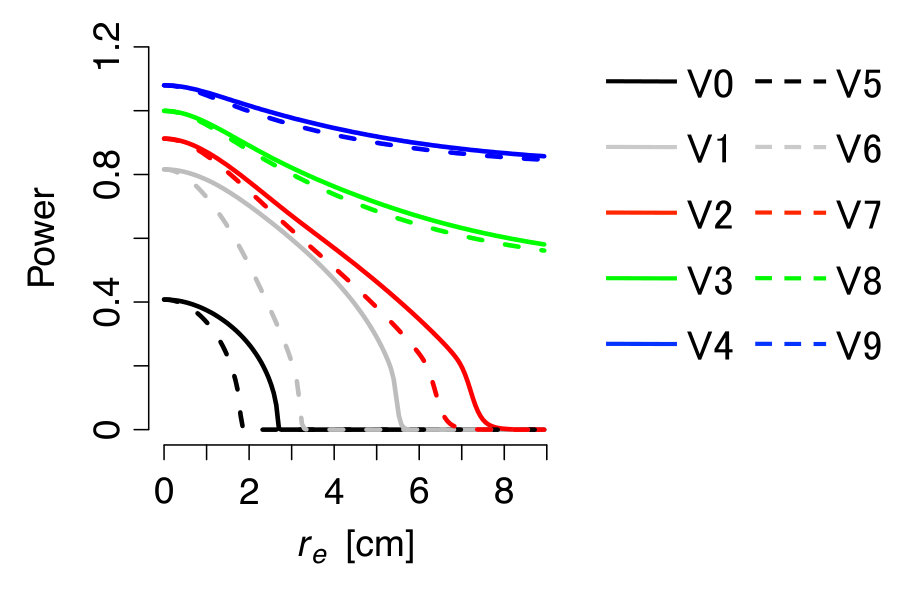 Figure 6
Figure 6| Parameter | |||||||
|---|---|---|---|---|---|---|---|
| Group S | -0.1 | -0.5 | 10msec | 80msec | -0.1 | 24rad/sec | 0.116 |
| Group H | -0.4 | -0.5 | 10msec | 80msec | -0.1 | 24rad/sec | -0.184 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Suppression of macroscopic oscillations in mixed populations of active and inactive oscillators coupled through lattice Laplacian
Ikuhiro Yamaguchi
Graduate School of Education, The University of Tokyo, 7-3-1 Hongo, Bunkyo-ku, Tokyo 113-0033, Japan
Takuya Isomura
Graduate School of Frontier Science, The University of Tokyo, 5–1–5 Kashiwanoha, Kashiwashi, Chiba 277–8563, Japan
Hiroya Nakao
Graduate School of Information Science and Engineering, Tokyo Institute of Technology, Tokyo 152-8522, Japan
Yutaro Ogawa
Graduate School of Frontier Science, The University of Tokyo, 5–1–5 Kashiwanoha, Kashiwashi, Chiba 277–8563, Japan
Yasuhiko Jimbo
Graduate School of Engineering, The University of Tokyo, 7-3-1, Hongo, Bunkyo-ku, Tokyo 113-8656, Japan
Kiyoshi Kotani
Research Center for Advanced Science and Technology, The University of Tokyo, 4-6-1, Komaba, Meguro-ku, Tokyo 153-8904, Japan
PRESTO, Japan Science and Technology Agency, 4-1-8, Honcho, Kawaguchi-shi, Saitama 332-0012, Japan
Abstract
We consider suppression of macroscopic synchronized oscillations in mixed populations of active and inactive oscillators with local diffusive coupling, described by a lattice complex Ginzburg-Landau model with discrete Laplacian in general dimensions. Approximate expression for the stability of the non-oscillatory stationary state is derived on the basis of the generalized free energy of the system. We show that an effective wavenumber of the system determined by the spatial arrangement of the active and inactive oscillators is an decisive factor in the suppression, in addition to the ratio of active population to inactive population and relative intensity of each population. The effectiveness of the proposed theory is illustrated with a cortico-thalamic model of epileptic seizures, where active and inactive oscillators correspond to epileptic foci and healthy cerebral cortex tissue, respectively.
coupled oscillators — aging transition — focal epilepsy
pacs:
05.45.Xt, 87.10.+e
I Introduction
Coupled-oscillator models have been widely used in the studies of a variety of rhythmic phenomena in physics, chemistry, and biology Kuramoto (2012); Winfree (2001); Ashwin et al. (2016). While many results for coupled oscillators have been obtained for phase models, in which the amplitude variable of oscillators is eliminated and only the phase of oscillators is considered, the effect of amplitude degrees of freedom often leads to intriguing collective dynamics that cannot occur in phase models.
Regarding collective oscillations in coupled oscillators with amplitude degrees of freedom, Daido and Nakanishi performed a pioneering study that revealed the effect of deteriorated inactive oscillators on macroscopic collective oscillation of the whole system, which they called aging transition Daido and Nakanishi (2004). In their model, macroscopic synchronized oscillation turns into a quiescent state as the proportion of inactive elements exceeds a certain critical value when coupling strength is sufficiently large.
The aging transition framework of Daido and Nakanishi, originally analyzed for globally coupled oscillators Daido and Nakanishi (2004); Pazó and Montbrió (2006); Tanaka et al. (2010); Daido (2011); Daido et al. (2013), has been generalized to various kinds of coupled-oscillator models with different coupling topologies. For example, aging transitions in a one-dimensional ring of locally coupled oscillators Daido (2008) and in networks of coupled oscillators with random coupling Morino et al. (2011); Tanaka et al. (2012) have been analyzed. However, compared with global coupling, mathematical treatment of other coupling topologies is difficult and relied mostly upon numerical simulations in most cases. As for coupled oscillators with lattice Laplacian coupling, which is a discretized version of ordinary diffusion in continuous media, spatial arrangement of the active and inactive oscillators decisively affects the aging transition in contrast to the case of global coupling.
The main aim of this paper is providing a theory to deal with heterogeneous spatial arrangement of active and inactive oscillators. We consider the suppression of macroscopic oscillations in mixed populations of active and inactive oscillators (i.e., aging transition) coupled through lattice Laplacian in general dimensions. By introducing a generalized free energy (Lyapunov function) of the system and approximating it with two variables, we show that an effective wavenumber of the system can be introduced, which characterizes the arrangement of the active and inactive oscillators, and can be used for determining whether suppression of macroscopic oscillation occurs or not.
Another aim of this paper is to show how the aging transition framework with the proposed theory provides insights into realistic situations in a cortico-thalamic model of epileptic seizures. Though the oscillating state and quiescent state were interpreted as alive and dead respectively in the original study Daido and Nakanishi (2004), the aging transition framework can also be interpreted in the opposite sense. That is, we consider whether pathological macroscopic oscillation caused by pathological oscillators can be suppressed when the proportion of the healthy oscillators is increased Kim et al. (2009). Using the proposed theory, we analyze a cortico-thalamic model of epileptic seizures, where active and inactive oscillators correspond to epileptic foci and healthy region, respectively.
II Theory
II.1 Model
We consider a mixed population of active and inactive oscillators, where the active group is denoted as S (seizure) and the inactive group is denoted as H (healthy). The oscillators are placed on a -dimensional hypercubic lattice of sites with a lattice constant , where is the side length of the hypercube, and periodic boundary conditions are assumed.
The system is described by the following lattice complex Ginzburg-Landau model on a -dimensional lattice ( is a natural number) of lattice constant :
[TABLE]
Here, is the complex amplitude of the oscillator at lattice coordinate and at time , is the natural frequency of the oscillator at the onset of oscillation, , and are real parameters, and represents a diffusion constant. The Hopf bifurcation parameter determines if the oscillator is active () or inactive (), and denotes the -dimensional discrete (lattice) Laplacian defined as
[TABLE]
For simplicity, we restrict our investigation to the case where all oscillators have the same frequency parameter . Then, by introducing a rotating frame with frequency and denoting , we obtain
[TABLE]
We also assume that the characteristics of the oscillators are binary. That is, if the oscillator belongs to group S, then its bifurcation parameter satisfies ; if the oscillator belongs to group H, . All parameters other than are identical in both groups.
Depending on the parameters and the spatial arrangement of the active and inactive oscillators, the system is in the quiescent state () or in the active state (). When the system is in the active state, that is, when the oscillators undergo collective oscillations, we can assume complete synchronization of the oscillators in each group because the frequency parameters of the oscillators are the same and the diffusion constant is real. That is, we suppose that the oscillation amplitudes of the oscillators are uniform in each group, i.e., in group S and in group H, where and are real amplitudes and the real-valued function represents the phase of the oscillation in the rotating frame. Namely, we assume that the oscillators are approximately binarized into either of the two groups. This binarization assumption holds strictly when is infinitesimal or when the oscillators of group S and H are arranged in the smallest possible checkered pattern. Though some error may generally be expected, this assumption also works reasonably well for other arrangements and makes the problem analytically tractable.
II.2 Two-variable free energy
It is known that Eq. (3) has a Lyapunov function, or the generalized free energy (GFE), which monotonously decreases with the evolution of the system Aranson and Kramer (2002). As we derive below, we can approximate the GFE of Eq. (3) by a two-variable function as
[TABLE]
under the two-group approximation that we introduce below, where represents th-order terms in and . Here, and are the proportions of the oscillators in group S and group H, respectively (), and the parameter represents the effective wavenumber characterizing the spatial arrangement of group S and H defined below.
The approximate GFE Eq. (4) can be obtained as follows. The exact GFE of Eq. (3) is given by Aranson and Kramer (2002)
[TABLE]
where denotes the discrete gradient operator on the -dimensional lattice defined as
[TABLE]
Using the above GFE, Eq. (3) and its complex conjugate can be expressed as
[TABLE]
where and . Note that is monotonically decreasing:
[TABLE]
where is used. Using the binarization assumption, the summation of the square amplitude in Eq. (6) can be approximated as
[TABLE]
which we call the two-group approximation.
To rewrite the summation of the discrete Laplacian term in Eq. (6), we use the discrete Fourier transformation (DFT),
[TABLE]
We can then rewrite the summation in Eq. (6) as
[TABLE]
where
[TABLE]
This expression for can be derived from the definition of the discrete gradient Eq. (7) as
[TABLE]
for the first component, and similarly for the other components. Note that when , and also that the difference between and is not negligible when is lager than or comparable to .
We now introduce an effective wavenumer :
[TABLE]
which characterizes the spatial arrangement of the active and inactive oscillators. The second expression follows from the approximation that the oscillators are binarized into two classes depending on the sign of . Plugging this definition into Eq. (12) and using Parseval’s identity, the summation can be approximated as
[TABLE]
where denotes the direct-current component of in the DFT under the binarization assumption, i.e., the area average of . Substituting Eq. (10) and Eq. (16) into Eq. (6), we obtain Eq. (4).
II.3 Stability condition
Using the approximate GFE in Eq. (4), we can derive the approximate conditions for the linear stability of the stationary quiescent state of the system. The Hessian matrix of Eq. (4) at the quiescent state is given by
[TABLE]
Thus, the quiescent state is linearly stable if and only if and , which results in the following two inequalities:
[TABLE]
The first inequality indicates that the average value of should be negative for the linear stability of the system. If the average value of is positive, the macroscopic oscillation cannot be suppressed by any . This inequality is interpreted as the condition that the inactive group wins over the active group when becomes infinitely large.
The second inequality shows that larger diffusion constant is required to suppress the macroscopic oscillation when the effective wavelength of the arrangement of active and inactive oscillators is larger (i.e., when the effective wavenumber is smaller). It is remarkable that both inequalities do not depend on and therefore they are valid in any dimensions.
III Application to the cortico-thalamic model
To illustrate how our approximate theory can provide insights into realistic problems, we consider the cortico-thalamic model proposed by Kim and Robinson Kim and Robinson (2007); Kim et al. (2009),
[TABLE]
where the field variable represents a mean firing rate of the neurons within each local area of the cortex, is a delayed value of (where is the time delay), parameterizes the strength of cortico-cortical activities, characterizes cortico-thalamic feedback, and controls the nonlinear term that takes into account the characteristics of neuronal firing.
The Kim-Robinson model is categorized into a neural field model, which describes how a mesoscopic quantity characterizing neural activity evolves over both space and time Pinotsis et al. (2014). This model undergoes Hopf bifurcation and exhibits collective oscillations, which is interpreted as the onset of an epileptic seizure Kim et al. (2009). Though Eq. (19) has a simple expression, it is actually a system with time delay and exhibits a wide variety of oscillatory dynamics. It is known that the model can reproduce many of the typical oscillatory waveforms observed in real electroencephalograms. By the center manifold reduction, the lattice complex Ginzburg-Landau model, Eq. (3), can be derived from Eq. (19) after spatial discretization (see Ref. Yamaguchi et al. (2011) for details). In the following, we consider Eq. (3) with the spatial arrangements of active and inactive oscillators assumed in Ref. Kim et al. (2009) and analyze the transition to macroscopic oscillations.
In Ref. Kim et al. (2009), the authors analyzed focal epilepsy both numerically and analytically using the model Eq. (19) by assuming that the parameters and are in the unstable regime (i.e., the field variable undergoes oscillations) in the epileptic foci of the cortex, while and are in the stable regime ( tends to vanish) in the remaining tissue. Their study implied that areas surrounding the epileptic foci play an essential role in the suppression of epileptic seizures. However, the analytical condition derived in Ref. Kim et al. (2009) was based on simplified assumptions and therefore not general enough.
In Ref. Kim et al. (2009), it is assumed that the inactive region (inactive oscillators in our discrete model) never oscillates at all, there is only a single active area and, the oscillation decays at the edge of the boundary. These assumptions lead to the prediction that the active region can be suppressed whenever the diffusion constant is larger than a critical value. This prediction, however, is valid only in special cases. Consider the limiting case where the proportion of the active area is close to , where the activity of the active area is infinitely large, where the inactive area is vanishingly small, or where the inactivity (resistance to activation) of the inactive area is infinitely small. In these cases, it is obvious that the active region would not be suppressed no matter how large the diffusion constant is. As we show below, not the inactive group but the active group wins under sufficiently strong coupling in these cases.
The major difficulty in deriving a better analytical prediction is that Eq. (19) has a delay term, which complicates theoretical analysis. This difficulty can be circumvented by reducing Eq. (19) with time delay to the lattice complex Ginzburg-Landau model Eq. (3) by the center manifold reduction near the bifurcation point, as we considered in Ref. Yamaguchi et al. (2011). In the reduced Eq. (3), epileptic foci and areas surrounding them correspond to the regions with and , respectively. See Ref. Yamaguchi et al. (2011) for further details on Eq. (19) and its reduction to Eq. (3). The other problem is that the stability condition for mixed populations of coupled active and inactive oscillators on a two-dimensional lattice is difficult to derive analytically. By using the approximate theory that we developed in the previous section, we can analyze the effect of spatial arrangement of the active and inactive oscillators, extending the result of Ref. Kim et al. (2009).
In the following, we illustrate our theory by numerically integrating Eq. (19), which is spatially discretized on a 6 6 lattice, for several spatial arrangements of the oscillators.
III.0.1 Investigated patterns
Figure 1 shows the investigated arrangements of the active and inactive oscillators. The yellow cells represent active oscillators in group S, while the white cells represent inactive oscillators in group H. We study the following two series of spatial arrangements: K-series (K0, K1, …, K9) and V-series (V0, V1, …, V9). The pattern identifier is indicated to the left of each pattern. The value below the identifier represents the effective wavenumber multiplied by the side length, , defined by Eq. (15). In the K-series, = 4/36 is fixed and all the patterns satisfy the first inequality of Eq. (18), while in the V-series, varies from 4/36 to 28/36 and V3, V4, V8 and V9 do not satisfy the first inequality of Eq. (18).
III.0.2 Results for the K-series
Using our approximate theory, we first reproduce the typical results presented in Ref. Kim et al. (2009). Here, (= 60 cm) corresponds to the side length of a square of the cortex. There are four active oscillators at the center of the square as in the pattern K0 in Fig.1, which correspond to the focal epileptic area of the cortex. The parameter values used in our calculations are shown Table 1, which are the same as those used in Ref. Kim et al. (2009).
In Ref. Kim et al. (2009), it is shown that the macroscopic oscillation, which is considered to be a seizure, does not occur if is larger than about 2.5 cm in this case. This suggests that inhibition by the surrounding area of the epileptic focus plays an important role in the suppression of epileptic seizures.
Steady states of the system after a sufficiently long transient period are shown in Fig. 2 for several values of . The top row shows peak to peak amplitude of the oscillators. The waveforms of all oscillators are shown in the bottom row of Fig. 2. When , the oscillators are completely separated into two groups, i.e., oscillating group and quiescent group (the waveform of the quiescent group lies on the horizontal axis). As becomes larger, the inactive oscillators start to oscillate, while the active oscillators start to decrease their oscillation amplitudes, and all oscillators stop at the critical value of . We evaluated the critical value of the diffusion constant from the stability condition given by Eq. (18) and obtained 2.51 cm for the pattern K0. We can confirm that the oscillations actually disappear near this predicted point, in good agreement with Ref. Kim et al. (2009).
Figure 3 shows the results for all K-series patterns in Fig.1 in grayscale, and Fig. 4 shows the power of the macroscopic oscillation vs. , where the power is calculated as the areal and temporal mean square of the amplitudes of all oscillators. The oscillations disappear near the critical diffusion constants predicted by the present approximate theory.
III.0.3 Results for the V-series
Figures 5 and 6 show the results for the V-series patterns in Fig. 1. In these cases, the oscillations should remain at any large in V3, V4, V8 and V9, because the first inequality of Eq. (18) is not satisfied in these patterns. The validity of the prediction can be seen in Fig. 6. As for the other patterns, we can again confirm that the oscillations disappear near the critical diffusion constants predicted by the second inequality of Eq. (18).
The prediction errors are less than about 30% of the true values through K-series and V-series. These errors are caused by the fact that the binarization assumption does not hold exactly, especially when the characteristic length scale of the patterns is large. But the error is still not fatal and does not undermine the insight; we can still predict whether the suppression of the macroscopic oscillation occurs or not by the approximate theory.
IV DISCUSSION AND CONCLUSION
We have derived an approximate condition for the suppression of macroscopic oscillations in mixed populations of coupled active and inactive oscillators using the two-group approximation for the lattice complex Ginzburg-Landau model, which can be considered an aging transition due to local diffusive coupling. As illustrated by numerical simulations, the approximate condition, Eq. (18), explains the transitions qualitatively well. Although quantitative accuracy is not very well and further improvement in the approximation is desirable, we believe that the theory developed in this paper provides an important step toward understanding of the aging transition in mixed populations coupled oscillators.
In the present study, we focused our attention on the generalized free energy (GFE) and approximated it to analyze the transition in a simple way. Although the approximate GFE does not have the same information as the original dynamical equation in the sense that we cannot derive the original equation from the approximate GFE, it contains essential information for predicting the stability and helps us derive the stability condition. Note that, unlike the real Ginzburg-Landau equation, several conditions must be satisfied for the complex Ginzburg-Landau equation to possess a GFE function.
In our analysis, Fourier transform to the wavenumber space was used to extract the effective wave number of the spatial arrangement of the mixed oscillator population, which played an important role in determining the stability of the quiescent state in addition to the proportions of the oscillators () and the bifurcation parameters (). The following proposition is useful in understanding the meaning of and in calculating its actual value: the binding number between group S and group H is related to the effective wavenumber as . We can prove this proposition by noting that takes a non-zero value only at the boundaries between the group S and group H under our binarization assumption. Plugging at the boundaries and otherwise into Eq. (16) leads
[TABLE]
which gives , using .
Finally, though we illustrated the theory for a model of epileptic seizures, we stress that our result is generally applicable to coupled-oscillator models near the Hopf bifurcation, because the complex Ginzburg-Landau equation is a normal form of coupled oscillators near the Hopf bifurcation derived by the center manifold reduction method from the original model Yamaguchi et al. (2011); Yang and Robinson (2017).
Acknowledgements.
The authors would like to thank Yoshiharu Yamamoto, Toru Nakamura, Fumiharu Togo, Akifumi Kishi and Jerome Foo for useful discussion. This work is partly supported by JSPS KAKENHI Grant number 15K01499 and 18K17887 to IY.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kuramoto (2012) Y. Kuramoto, Chemical oscillations, waves, and turbulence , Vol. 19 (Springer Science & Business Media, 2012).
- 2Winfree (2001) A. T. Winfree, The geometry of biological time , Vol. 12 (Springer Science & Business Media, 2001).
- 3Ashwin et al. (2016) P. Ashwin, S. Coombes, and R. Nicks, The Journal of Mathematical Neuroscience 6 , 1 (2016).
- 4Daido and Nakanishi (2004) H. Daido and K. Nakanishi, Physical review letters 93 , 104101 (2004).
- 5Pazó and Montbrió (2006) D. Pazó and E. Montbrió, Physical Review E 73 , 055202 (2006).
- 6Tanaka et al. (2010) G. Tanaka, Y. Okada, and K. Aihara, Physical Review E 82 , 035202 (2010).
- 7Daido (2011) H. Daido, Physical Review E 84 , 016215 (2011).
- 8Daido et al. (2013) H. Daido, A. Kasama, and K. Nishio, Physical Review E 88 , 052907 (2013).