Can a flux-based mechanism explain positioning of protein clusters in a three-dimensional cell geometry?
Matthias Kober, Silke Bergeler, Erwin Frey

TL;DR
This study introduces a three-dimensional mathematical model demonstrating that flux-based mechanisms can effectively position protein clusters in bacterial cells, accounting for realistic cell geometries and diffusion processes.
Contribution
The paper extends previous flux-based positioning models to three-dimensional cell geometries, showing that cluster localization is robust in more realistic biological contexts.
Findings
Cluster moves to midcell in 3D geometry
Diffusion in cytosol is crucial for positioning
Mechanism positions two clusters equidistantly
Abstract
The plane of bacterial cell division must be precisely positioned. In the bacterium Myxococcus xanthus, the proteins PomX and PomY form a large cluster, which is tethered to the nucleoid by the ATPase PomZ and moves in a stochastic, but biased manner towards midcell, where it initiates cell division. Previously, a positioning mechanism based on the fluxes of PomZ on the nucleoid was proposed. However, the cluster dynamics was analyzed in a reduced, one-dimensional geometry. Here we introduce a mathematical model that accounts for the three-dimensional shape of the nucleoid, such that nucleoid-bound PomZ dimers can diffuse past the cluster without interacting with it. Using stochastic simulations, we find that the cluster still moves to and localizes at midcell. Redistribution of PomZ by diffusion in the cytosol is essential for this cluster dynamics. Our mechanism also positions two…
Click any figure to enlarge with its caption.
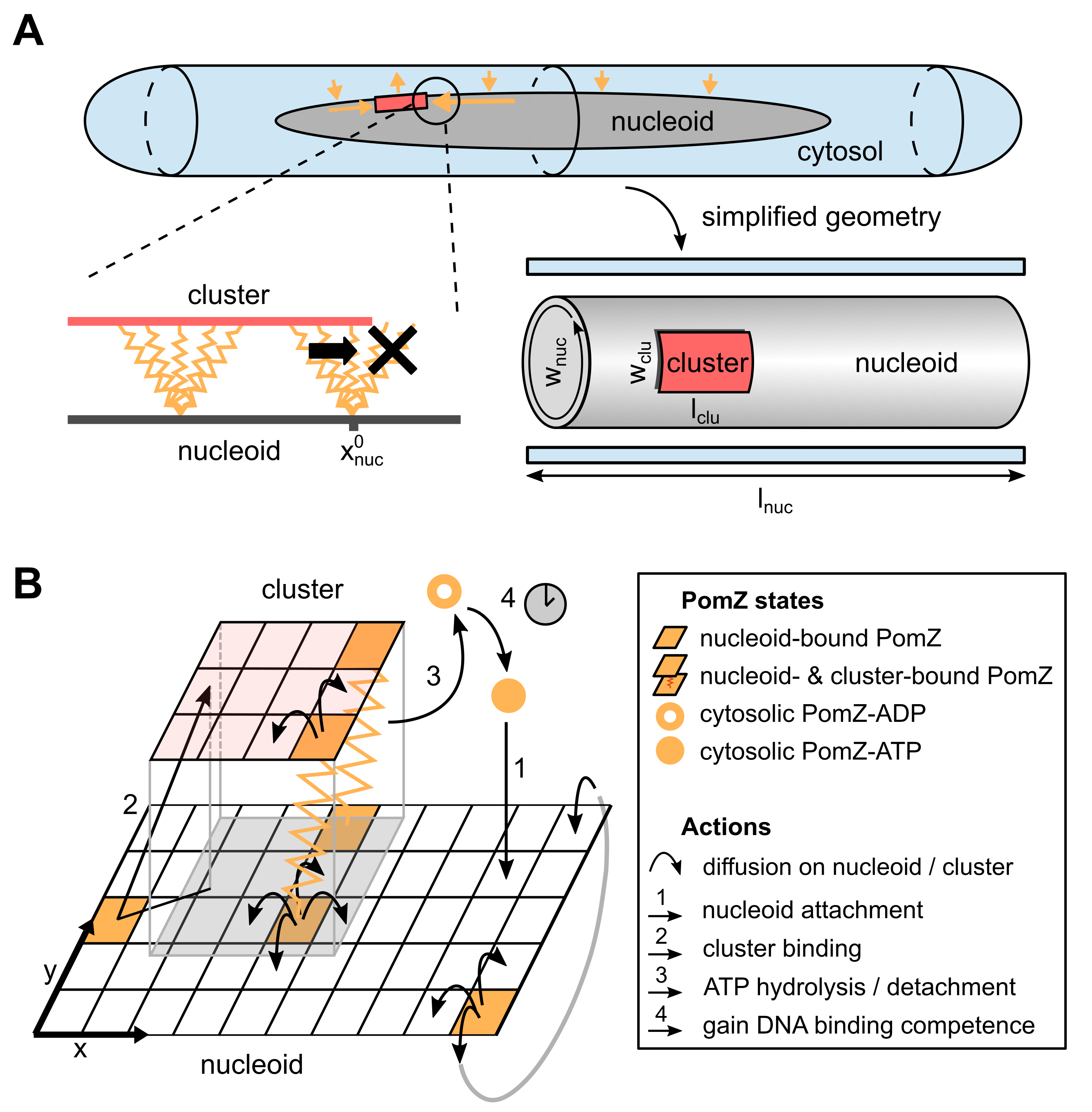 Figure 1
Figure 1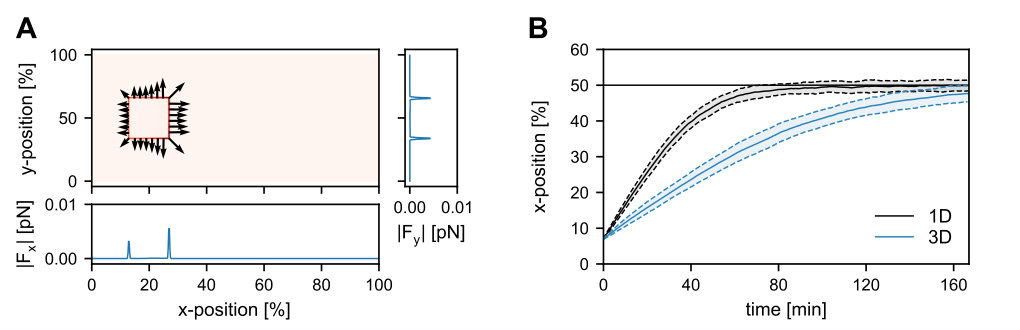 Figure 2
Figure 2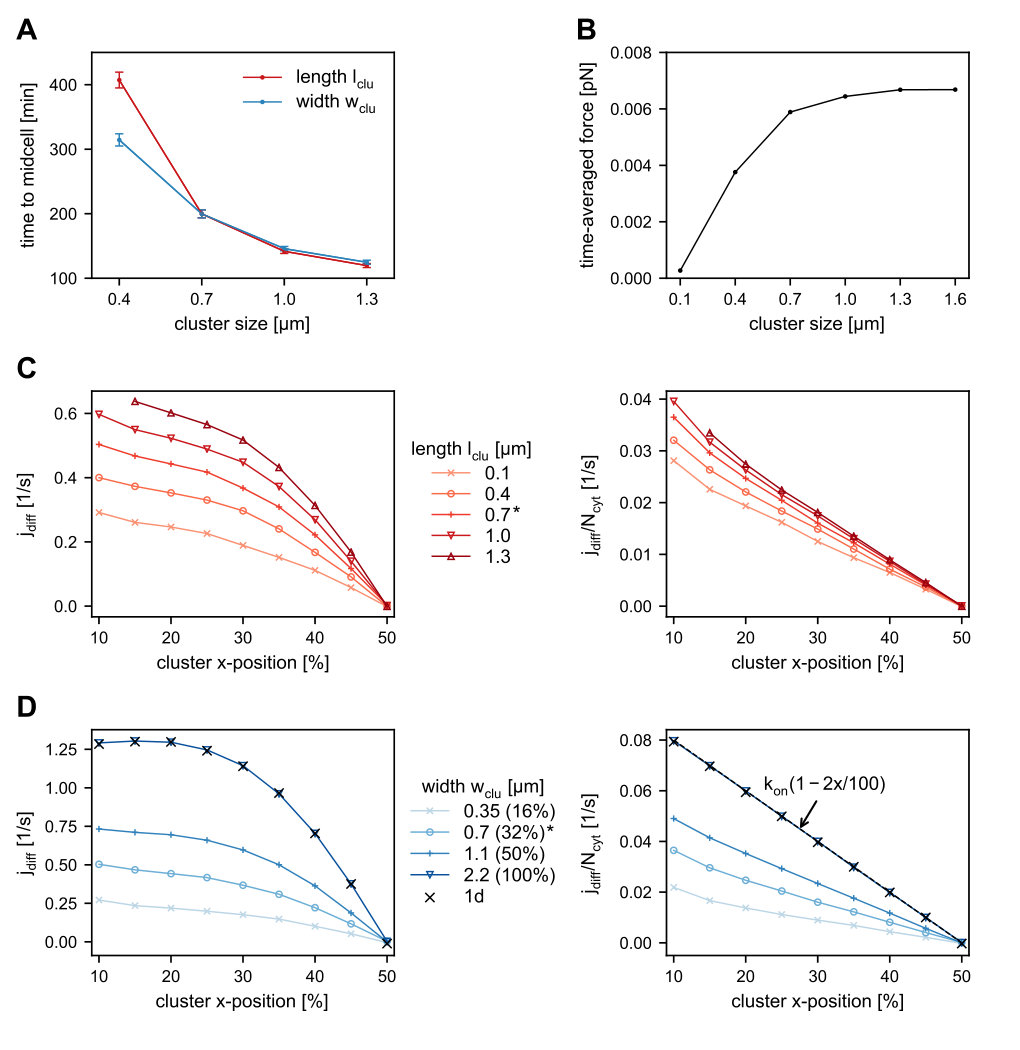 Figure 3
Figure 3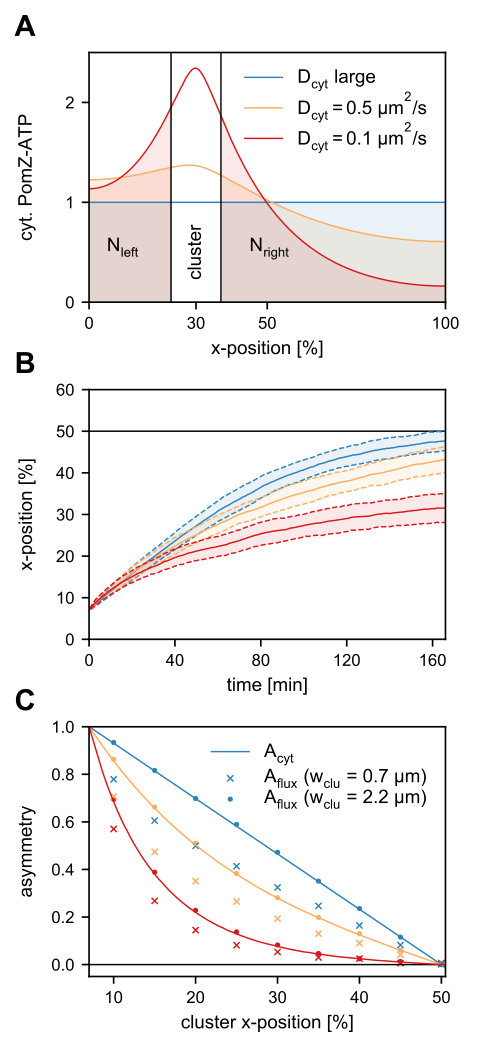 Figure 4
Figure 4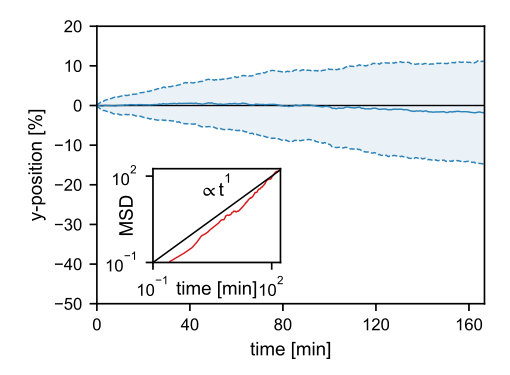 Figure 5
Figure 5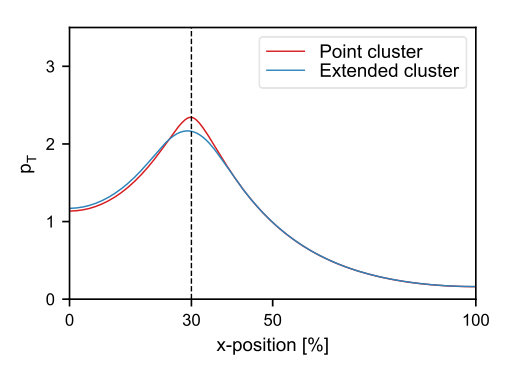 Figure 6
Figure 6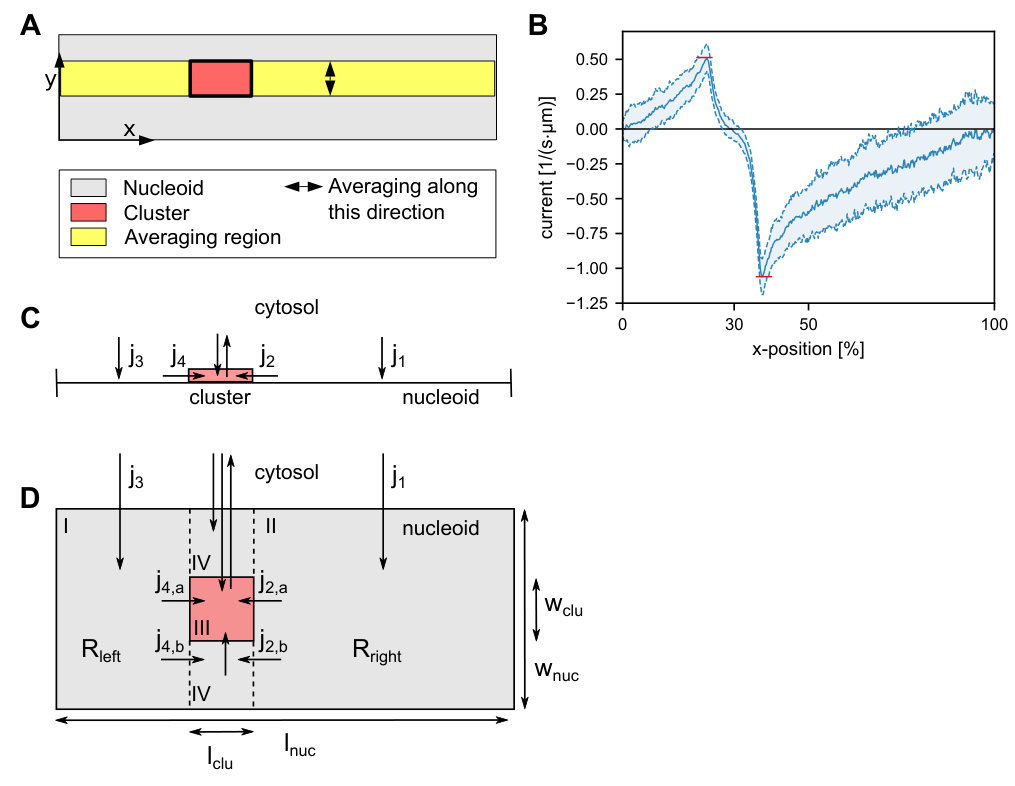 Figure 7
Figure 7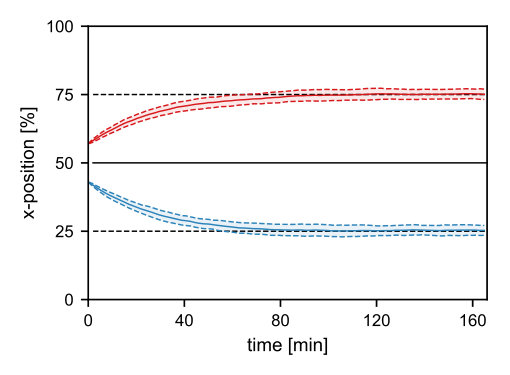 Figure 8
Figure 8Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
††thanks: Correspondence: [email protected]
Can a flux-based mechanism explain positioning of protein clusters in a three-dimensional cell geometry?
Matthias Kober
These two authors contributed equally
Silke Bergeler
These two authors contributed equally
Erwin Frey
Arnold Sommerfeld Center for Theoretical Physics and Center for NanoScience, Department of Physics, Ludwig-Maximilians-Universität München, Theresienstraße 37, D-80333 Munich, Germany
Abstract
The plane of bacterial cell division must be precisely positioned. In the bacterium Myxococcus xanthus, the proteins PomX and PomY form a large cluster, which is tethered to the nucleoid by the ATPase PomZ and moves in a stochastic, but biased manner towards midcell, where it initiates cell division. Previously, a positioning mechanism based on the fluxes of PomZ on the nucleoid was proposed. However, the cluster dynamics was analyzed in a reduced, one-dimensional geometry. Here we introduce a mathematical model that accounts for the three-dimensional shape of the nucleoid, such that nucleoid-bound PomZ dimers can diffuse past the cluster without interacting with it. Using stochastic simulations, we find that the cluster still moves to and localizes at midcell. Redistribution of PomZ by diffusion in the cytosol is essential for this cluster dynamics. Our mechanism also positions two clusters equidistantly on the nucleoid. We conclude that a flux-based mechanism allows for cluster positioning in a biologically realistic three-dimensional cell geometry.
Keywords
midcell localization, Myxococcus xanthus, cell division, stochastic simulation,
flux-based mechanism
I Introduction
In bacteria, intracellular positioning of proteins is important for vital biological processes, including localization of the cell division machinery to midcell, as well as chromosome and plasmid segregation. The positioning systems responsible often involve P-loop ATPases such as ParA and MinD Gerdes2010 ; Lutkenhaus2012 . These ATPases switch between an ATP- and ADP-bound state, which alters their subcellular localization: the ATP-bound form typically binds as a dimer to the nucleoid or membrane, while the ADP-bound form diffuses freely in the cytosol. Activating proteins stimulate the ATPase activity of the ParA-like proteins, which results in detachment of the protein from its respective scaffold in the ADP-bound form. Intracellular patterns of these ParA-like ATPases depend on the binding properties of the ADP- and ATP-bound forms, the localization of the stimulating proteins, and the cell geometry Thalmeier2016 .
Recently, a protein system that includes a ParA-like ATPase has been identified, which regulates the localization of the cell division site in the bacterium Myxococcus xanthus Schumacher2017a ; Treuner-Lange2013 ; Schumacher2017 . The Pom system consists of three proteins: PomX, PomY and PomZ. The ATP-bound ParA-like ATPase PomZ binds as a dimer non-specifically to DNA. PomX and PomY form a macromolecular cluster, the PomXY cluster, which is tethered to the nucleoid via PomZ dimers. This tethering is transient, since PomX, PomY and DNA synergistically stimulate the ATPase activity of PomZ, which results in two ADP-bound PomZ monomers being released into the cytosol. Fluorescence labeling shows that, shortly after cell division, the PomXY cluster moves from a position close to one nucleoid end towards mid-nucleoid in a biased random walk-like movement that depends on PomZ. Once at mid-nucleoid, the Pom cluster positively regulates FtsZ ring formation Schumacher2017a .
Though the Pom system in M. xanthus and the Min system in Escherichia coli serve the same function (localization of the cell division plane), they differ in important respects: Dimeric MinD-ATP binds to the cell membrane, and MinC inhibits FtsZ ring formation outside of the midcell region, leading to negative regulation of mid-plane localization. In this regard, the Pom system is more similar to the Par system for low-copy-number plasmid and chromosome segregation, and related systems, e.g. those used to position chemotaxis protein clusters Roberts2012 and carboxysomes Savage2010 . These systems utilize ParA-like ATPases that bind in the ATP-bound form to the nucleoid and regulate the movement of a cargo (e.g. protein cluster, partition complex or plasmids) to the required intracellular positions Howard2010 .
Previously, we introduced a one-dimensional mathematical model of the dynamics of the PomXY cluster in M. xanthus Schumacher2017a ; Bergeler2018 that takes the elasticity of the nucleoid Wiggins2010 ; Lim2014 into account. Based on this model, we proposed a mechanism that relies on PomZ dimer fluxes on the nucleoid to generate the experimentally observed cluster dynamics, which is midcell localization. A flux-based mechanism was originally proposed by Ietswaart et al. Ietswaart2014 for Par-mediated plasmid positioning. They showed that diffusive ParA fluxes can in principle explain equidistant spacing of plasmids along the nucleoid, because only when this is the case will the fluxes from either side of each plasmid balance up. Hence, macromolecular objects can be evenly distributed on the nucleoid if they move in the direction from which the larger number of ParA proteins impinges upon them Ietswaart2014 . In our model of the Pom system, an analogous mechanism can localize the PomXY cluster to mid-nucleoid Schumacher2017a ; Bergeler2018 .
It remained unclear, however, whether such a flux-based mechanism can also localize midcell if the full three-dimensional geometry of the nucleoid is accounted for (see Figure 1A). The reason is that, in contrast to the one-dimensional case, PomZ dimers can now more easily pass the cluster by without interacting with it. As a result, an asymmetry in the PomZ fluxes might be balanced. Here we show that the Pom cluster still localizes at midcell by a flux-based mechanism even if PomZ dimers can diffuse past the cluster.
II Results
II.1 Flux-based mechanism for midcell localization
The dynamics of the PomXY cluster on the nucleoid crucially depends on the PomZ dynamics, as the cluster is tethered to the nucleoid via PomZ dimers Schumacher2017a . Based on the biochemical processes suggested by experiments Schumacher2017a , we model the dynamics of PomZ as follows (see Figure 1B and SI text for details). ATP-bound PomZ dimers bind to the nucleoid with rate (action 1 in Figure 1B). Once on the nucleoid, they diffuse with diffusion constant . The PomZ dimers are modeled effectively as springs to account for the elasticity of the chromosome and the Pom proteins (as in Lim2014 ; Surovtsev2016a ). For simplicity we will refer to the PomZ dimers as springs in the following, although the elasticity mainly originates from the nucleoid.
A nucleoid-bound PomZ dimer can attach to the cluster with rate either ‘orthogonally’, or obliquely in an extended configuration (action 2 in Figure 1B). We assume that cluster-bound PomZ can diffuse on both the nucleoid and the PomXY cluster. However, the freedom of movement of nucleoid- and cluster-bound PomZ is restricted due to the energy cost involved in stretching the spring (for details see SI text). Cluster-bound PomZ dimers remain attached to the cluster until they are released into the cytosol upon ATP hydrolysis, which is stimulated by PomX, PomY and DNA. ATP hydrolysis leads to a conformational change in the PomZ dimer and triggers the release of two ADP-bound PomZ monomers into the cytosol Schumacher2017a . In our model, we combine these processes into one by using a single rate to describe the detachment of cluster-bound PomZ-ATP dimers into the cytosol as monomers (action 3 in Figure 1B). Before PomZ can rebind to the nucleoid, it must first bind ATP and dimerize. This introduces a delay between detachment from and reattachment to the nucleoid, which allows for spatial redistribution of the quickly diffusing cytosolic PomZ dimers in the cell (action 4 in Figure 1B).
The cluster dynamics, which we approximate as over-damped, is determined by the forces exerted by the PomZ dimers on the cluster and the friction coefficient of the cluster (see SI text). Previously we suggested a flux-based mechanism for its midcell positioning in a one-dimensional model geometry Schumacher2017a ; Bergeler2018 , which can be summarized as follows. The PomZ dimers, which are modeled as springs, can exert net forces on the cluster in two different ways. They can attach to the cluster in a stretched configuration and thereby exert forces on the cluster (similar to the DNA-relay mechanism proposed for Par systems, Lim2014 ). However, for PomZ dimers that quickly diffuse on the nucleoid (as observed experimentally in M. xanthus cells) the initial deflection of the spring relaxes within a short time, such that only a small net force is exerted on the cluster. In this case, forces are mainly generated by cluster-bound PomZ dimers that encounter the cluster’s edge (Figure 1A, bottom left) Bergeler2018 . Every time a nucleoid- and cluster-bound PomZ dimer reaches the cluster’s edge, the nucleoid binding site can diffuse beyond the cluster region, whereas the cluster binding site is restricted in its movement, which on average results in a net force on the cluster. These forces are such that a protein that reaches the cluster from the right exerts a force, which ‘drags’ the cluster to the right and vice versa.
The fact that PomZ dimers can exert forces on the cluster is not enough to explain the movement of the cluster towards midcell. Here, the asymmetry in the PomZ dimer density on the nucleoid, which prevails as long as the cluster is located off center, i.e. not already at mid-nucleoid, is crucial. Since PomZ dimers can attach anywhere on the nucleoid but can detach only when they make contact with the cluster (as stimulation of their ATPase activity depends on PomX, PomY and DNA), a non-equilibrium flux of PomZ dimers in the system can be maintained. In particular, this flux includes a diffusive flux of PomZ dimers along the nucleoid towards the cluster. For a cluster located off-center the diffusive PomZ fluxes into the cluster from either side will differ Ietswaart2014 . Since the average forces applied by PomZ dimers arriving from the right and left sides of the cluster act in opposite directions, the difference in the PomZ fluxes from the two sides determines the net force exerted on the cluster Bergeler2018 . Taken together, these factors combine to drive a self-regulated midcell localization process as long as the PomZ dynamics is fast compared to the cluster dynamics and, if this is not the case, lead to oscillatory cluster movements along the nucleoid Bergeler2018 .
II.2 A three-dimensional model for midcell localization
To understand how the geometry of the nucleoid and the size of the PomXY cluster affect the cluster dynamics and thereby test whether a flux-based mechanism is feasible in a biologically realistic three-dimensional geometry, we investigated the mathematical model illustrated in Figure 1. Here, the nucleoid and the PomXY cluster are approximated as a cylindrical object and a rectangular sheet, respectively. Since experiments in M. xanthus cells suggest that the cluster is large Schumacher2017a we assume that the cluster, tethered to the nucleoid via PomZ dimers, moves over the nucleoid’s surface and does not penetrate the bulk of the nucleoid. Moreover, we assume that PomZ dimers also bind to and diffuse on the nucleoid’s surface only.
The cylindrical geometry of the nucleoid is mathematically implemented by a rectangular sheet with periodic boundary conditions for the PomZ movements along the short cell axis ( direction) and reflecting boundary conditions along the long cell axis ( direction, Figure 1B). The cluster is modeled as a rectangle with reflecting boundaries at its edges for the PomZ dimer movement. We refer to the extension of the cluster along the long and short cell axis as the cluster’s length, , and width, , respectively (see Figure 1A).
Besides the nucleoid and the cluster, the cytosol needs to be accounted for in our model, as PomZ dimers cycle between a nucleoid-bound and cytosolic state. We expect the cytosolic diffusion constant of PomZ to be of the same order as that of MinD proteins in E. coli cells, 10\text{,}\mathrm{\SIUnitSymbolMicro}\mathrm{m}^{2}\mathrm{s}^{-1} [Meacci2006 ](#bib.bib15). For ParA ATPases involved in chromosome and plasmid segregation, a delay between the release of ParA from the nucleoid into the cytosol and the re-acquisition of its capacity for non-specific DNA binding is observed experimentally [Vecchiarelli2010 ](#bib.bib16). Upon ATP hydrolysis and release of two ADP-bound ParA monomers into the cytosol, ParA must bind ATP, dimerize and then regain the competence to bind non-specifically to DNA before it can reattach to the nucleoid. The last step, a conformational change in the ATP-bound ParA dimer, is the slowest of these processes, occurring on a time scale of the order of minutes, $\tau\approx$5\text{\,}\min Vecchiarelli2010 . Since PomZ is ParA-like, we expect the PomZ ATPase cycle to be similar to the ParA cycle, and therefore assume the corresponding reaction rates to be of the same order of magnitude. This yields the following estimate for the diffusive length of cytosolic PomZ, 55\text{,}\mathrm{\SIUnitSymbolMicro m}$$, which is significantly larger than the average cell length of a M. xanthus cell of Schumacher2017a . Hence, the assumption of a well-mixed PomZ density in the cytosol is justified Thalmeier2016 .
However, it is not known how the cytosolic distribution of PomZ affects the cluster dynamics in our proposed flux-based mechanism. In particular, does a realistic, non-uniform distribution increase or reduce the speed of the cluster movement towards midcell? To obtain a qualitative answer to this question, we accounted for the cytosolic PomZ distribution in a simplified way by focusing on the variation in PomZ density along the long cell axis and approximating the density along the short cell axis as uniform (Figure 1A, bottom right). In section II.5 we discuss how the cluster dynamics is affected when PomZ’s diffusion constant in the cytosol is reduced (which leads to deviations from the uniform PomZ distribution), but for now we assume a homogeneous PomZ-ATP distribution in the cytosol.
II.3 A flux-based mechanism can explain midcell positioning in three dimensions
Our simulations using the three-dimensional model geometry show that the net force exerted by the PomZ dimers on a stationary cluster (i.e. a cluster with fixed position) is still directed towards mid-nucleoid (Figure 2A). The asymmetry in the forces can be attributed to an asymmetry in the PomZ fluxes along the nucleoid into the cluster. This finding indicates that the cluster movement is biased towards midcell in the three-dimensional model geometry also. Hence, a flux-based mechanism is also conceivable in three dimensions. Indeed, the simulated PomXY cluster trajectories show movement towards and localization at mid-nucleoid (Figure 2B). If the cluster’s width does not cover the complete nucleoid circumference, the movement towards midcell takes longer than in the one-dimensional case (Figure 2B). The forces along the short cell axis direction balance (Figure 2A) and hence no bias in the cluster movement along the short axis is expected for a slowly moving cluster and fast PomZ dynamics. The simulated cluster trajectories show diffusive motion along the short cell axis direction for the parameters given in Table S1 (see SI text).
II.4 The cluster’s linear dimensions determine the time taken to reach midcell
In this section we ask how the arrival time of the cluster at mid-nucleoid depends on the linear dimensions of the cluster. Our simulations show that the larger the cluster’s length or width, the faster the cluster moves towards midcell (Figure 3A). In the following we discuss how these two observations can be explained heuristically.
The net force applied to the cluster by PomZ dimers depends on the average force exerted by a single PomZ dimer and on the flux difference in PomZ dimers arriving at the cluster Bergeler2018 . The latter can be regarded as the frequency of PomZ interactions with the cluster that lead to a net force contribution.
The flux difference into the cluster increases for larger or broader clusters for two reasons. First, increasing the length or width of the cluster results in an increased number of cluster-bound PomZ dimers, as the cluster becomes more accessible to nucleoid-bound PomZ dimers. This increase in cluster-bound PomZ dimers leads to a larger turnover of PomZ dimers cycling between the nucleoid-bound and cytosolic state, which also increases the flux difference. Second, the larger and wider the cluster, the less likely it is that PomZ dimers will diffuse past it without attaching, which would otherwise reduce the flux difference into the cluster along the long cell axis.
The forces exerted by PomZ dimers also depend on the linear dimensions of the cluster. Increasing the cluster size increases the average force exerted by a single PomZ dimer, until a maximal force is reached (Figure 3B). This dependence can be explained by diffusion of PomZ dimers bound to the cluster: The smaller the cluster size, the more likely it is that a PomZ dimer attaching close to one edge of the cluster will reach the other edge before detaching into the cytosol. Since the PomZ dimer then exerts forces in both directions along the long cell axis, the time-averaged force it applies to the cluster is reduced.
Based on these observations, we can rationalize the dependence of the arrival time at mid-nucleoid on the length of the cluster as follows. An increase in cluster length (while keeping its width constant) increases the numbers of PomZ dimers interacting with the cluster, and hence the flux difference along the long cell axis. Indeed, our simulation results show that the longer the cluster, the larger the difference in the PomZ dimer fluxes from either side along the direction of the long cell axis (Figure 3C). To test whether the increased flux of PomZ dimers in the system is the main determinant for the observed increase in the flux difference, we scaled the fluxes by the number of PomZ dimers in the cytosol, , which is proportional to the flux of PomZ dimers onto the nucleoid. Upon rescaling, the flux differences decay approximately linearly with the cluster position (Figure 3C, right). However, longer clusters still show the largest flux differences. This phenomenon can be attributed to the fact that for shorter clusters PomZ dimers are more likely to diffuse past the cluster.
For a longer cluster not only the diffusive flux of PomZ dimers into the cluster, but also the force exerted by a single PomZ dimer on the cluster is increased (Figure 3B). Hence, frequency and magnitude of forces exerted on the cluster are increased, implying a larger net force, which explains the shorter arrival times of the cluster at mid-nucleoid (Figure 3A).
Next, we discuss the dependence of the arrival time on the cluster width. As in the case of an increase in length, the overall turnover of PomZ dimers increases with cluster width. The width of the cluster determines how many PomZ dimers approach and attach to the cluster from the directions of the long and the short cell axes, respectively, and how many pass the cluster without interacting with it. The broader the cluster, the smaller the flux of PomZ dimers that can diffuse past the cluster without attaching. Our simulation results show an increased PomZ flux difference when the cluster width is increased from to of the nucleoid’s circumference (Figure 3D). When the cluster covers the entire circumference of the nucleoid, we call it a ring. Rescaling of the flux differences with the number of PomZ dimers in the cytosol, , leads to values that are still larger for wider clusters, due to the reduced flux of PomZ dimers past the cluster (Figure 3D, right). The forces exerted by single PomZ dimers in the direction of the long cell axis direction are not affected by a change in the cluster width. Hence, the decrease in arrival time for wider clusters can be explained by the increased PomZ flux difference into the cluster along the long cell axis alone.
II.5 Cytosolic diffusion ensures fast midcell positioning of the cluster
So far, we have assumed that the cytosolic PomZ distribution is spatially uniform. Now we investigate how the cluster dynamics change when spatial heterogeneity in the cytosolic PomZ distribution is included in the model. To this end, we explicitly incorporate the cytosol by approximating the cytosolic volume as a one-dimensional layer of the same length as the nucleoid (see Figure 1A) and formulate reaction-diffusion equations for the density of cytosolic PomZ-ADP () and PomZ-ATP () along the long cell axis. For simplicity, we consider only an active (ATP-bound) and inactive (ADP-bound) conformation of PomZ, and disregard any explicit monomeric and dimeric states of PomZ in the cytosol (for details see SI text). The stationary solution for deviates most from a uniform distribution, the smaller the cytosolic diffusion constant (Figure 4A). In the limit of infinitely large cytosolic PomZ diffusion constants, the cytosolic PomZ distribution becomes spatially uniform.
To investigate the effect of the cytosolic PomZ distribution on the cluster’s trajectory, we replaced the spatially uniform cytosolic PomZ-ATP distribution by , in our three-dimensional model. Since M. xanthus cells are rod-shaped with the length being much larger than the width, we approximate the cytosolic PomZ-ATP distribution along the short cell axis as uniform. With this spatially heterogeneous attachment rate to the nucleoid, our simulations show that for a larger deviation of the cytosolic PomZ distribution from a spatially uniform one (decreasing ), the movement of the clusters is less biased towards mid-nucleoid (Figure 4B). We conclude that the cytosolic PomZ distribution has an impact on the cluster trajectories and the velocity of the cluster towards mid-nucleoid is maximal for a uniform cytosolic PomZ-ATP distribution.
How does the cytosolic PomZ distribution affect the cluster’s movement? The flux of PomZ dimers onto the nucleoid region to the left of the cluster and the diffusive flux of PomZ dimers into the cluster from the left are equal for a stationary cluster in the one-dimensional model geometry, and vice versa for the right side (see SI text). Since the total flux of PomZ dimers onto the nucleoid to the left and right of the cluster depends on the cytosolic PomZ-ATP distribution, we expect the diffusive PomZ fluxes into the cluster to depend on this distribution as well. To investigate this further, we define the following asymmetry quantity
[TABLE]
which compares the total numbers of cytosolic PomZ-ATP left and right of the cluster (see Figure 4A):
[TABLE]
The corresponding asymmetry in the PomZ fluxes into the cluster from the left, , and right, , sides is given by
[TABLE]
We measured this flux asymmetry in our simulations for two scenarios. First, for a PomXY cluster that forms a ring around the nucleoid and second, for a cluster that does not entirely encompass the nucleoid’s circumference. We find that the asymmetry in the flux, , obtained from simulations for a cluster that forms a ring, and the corresponding asymmetry in the cytosolic PomZ-ATP density, , agree nicely (Figure 4C). Hence, an asymmetry in the cytosolic distribution of PomZ-ATP is directly reflected in the diffusive PomZ fluxes into the cluster. For an infinitely large cytosolic PomZ diffusion constant, both asymmetry measures decay linearly when the cluster position is varied from far off-center towards mid-nucleoid. Decreasing the cytosolic PomZ diffusion constant results in asymmetry curves that decay faster than linearly towards zero (Figure 4C). For a cluster that does not cover the whole nucleoid’s circumference, the asymmetry in the PomZ fluxes into the cluster is smaller than the asymmetry in the cytosolic PomZ-ATP concentration (Figure 4C). This can be attributed to the reduction in the diffusive fluxes of PomZ dimers into the cluster along the long cell axis, as discussed before. The reduced asymmetry in the diffusive PomZ fluxes explains the less biased movement of the cluster towards mid-nucleoid for smaller cytosolic PomZ diffusion constants (Figure 4B).
II.6 Two clusters localize at one- and three-quarter positions
Motivated by equidistant positioning of multiple cargoes, such as plasmids Ringgaard2009 ; Ebersbach2006 ; Sengupta2010 ; Ietswaart2014 ; Hatano2010 , we also considered the dynamics of two PomXY clusters in the three-dimensional model geometry. Our simulation results show equidistant positioning of the two clusters (see SI text, Figure S4).
III Discussion
In this work we investigated a mathematical model for midcell localization in M. xanthus using a biologically realistic three-dimensional geometry for the nucleoid. Whether or not a flux-based mechanism can position macromolecular objects when the ATPases (here PomZ) can diffuse past a cargo (here PomXY cluster) has been questioned, because the fluxes into the cargo might equalize Ietswaart2014 . We showed that if PomZ dimers can diffuse past the PomXY cluster, there is still a flux difference into the cluster (for a cluster positioned off-center), which leads to a bias in the cluster movement towards mid-nucleoid. Hence, we conclude that a flux-based mechanism can explain midcell positioning of one or equidistant localization of several Pom clusters even if PomZ can diffuse past the cluster.
To investigate the effect of the flux of PomZ dimers past the cluster, we studied the dependence of cluster dynamics on its width and length. We find that increasing the cluster length or width shortens the time taken for the cluster to reach mid-nucleoid. This can be attributed to an overall increase in the flux difference, reduced flux past the cluster and the larger forces single PomZ dimers exert, on average, on larger and wider clusters.
Our simulation data further demonstrates that fast cytosolic diffusion of PomZ proteins reduces the time taken to find midcell. This finding is in agreement with previous results for dynamic protein clusters in bacterial cells Murray2017 . The asymmetry in the cytosolic PomZ density left and right of the cluster along the long cell axis is reflected in the diffusive flux difference of PomZ dimers into the cluster, and thus influences the cluster dynamics. If PomZ diffuses quickly in the cytosol, the cytosolic PomZ-ATP distribution becomes spatially uniform. In this case, the flux of PomZ dimers onto the nucleoid scales with the length of the nucleoid regions left and right of the cluster. This results in the largest PomZ flux differences (for an off-center cluster) compared to spatially non-uniform cytosolic PomZ-ATP distributions. Interestingly, spatial redistribution of proteins in the cytosol is also found to be important for other pattern-forming systems, including Min protein pattern formation Halatek2018 ; Halatek2018b and ParA-mediated cargo movement Vecchiarelli2010 .
Le Gall et al. LeGall2016 have shown that partition complexes as well as plasmids move within the nucleoid volume. In contrast, based on the large size of the PomXY cluster, we assumed that the movement of the cluster, tethered to the nucleoid via PomZ dimers, is restricted to the surface of the nucleoid. To verify our assumption, the position of the cluster relative to the nucleoid needs to be measured in vivo using e.g. super-resolution microscopy. In addition, since PomZ dimers are much smaller than the PomXY cluster, they might be able to diffuse within the nucleoid volume even though the cluster does not. It would be interesting to investigate this aspect further.
In summary, we have shown that a flux-based mechanism can explain midcell localization of one, and equidistant positioning of two clusters in a model geometry that allows the ATPase PomZ to diffuse past the clusters on the nucleoid. This observation is also important for other positioning systems, such as the Par system, which equidistantly spaces low-copy-number plasmids along the nucleoid. Understanding the differences and similarities between these positioning systems will help us to understand the generic mechanisms underlying the localization patterns of cargoes inside the cell.
Acknowledgements
The authors thank Isabella Graf, Emanuel Reithmann, Christoph Brand, Dominik Schumacher and Lotte Søgaard-Andersen for helpful discussions. This research was supported by a DFG fellowship through the Graduate School of Quantitative Biosciences Munich, QBM (SB, EF), the Deutsche Forschungsgemeinschaft (DFG) via project P03 within the Transregio Collaborative Research Center (TRR 174) “Spatiotemporal Dynamics of Bacterial Cells” (SB, EF), and the German Excellence Initiative via the program “Nanosystems Initiative Munich” (EF).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1(1) K. Gerdes, M. Howard, and F. Szardenings, “Pushing and pulling in prokaryotic DNA segregation,” Cell , vol. 141, no. 6, pp. 927–942, 2010.
- 2(2) J. Lutkenhaus, “The Par A/Min D family puts things in their place,” Trends Microbiol , vol. 20, pp. 411–418, 2012.
- 3(3) D. Thalmeier, J. Halatek, and E. Frey, “Geometry-induced protein pattern formation,” Proc Natl Acad Sci U S A , vol. 113, no. 3, pp. 548–553, 2016.
- 4(4) D. Schumacher, S. Bergeler, A. Harms, J. Vonck, S. Huneke-Vogt, E. Frey, and L. Søgaard-Andersen, “The Pom XYZ proteins self-organize on the bacterial nucleoid to stimulate cell division.,” Dev Cell , vol. 41, pp. 299–314, 2017.
- 5(5) A. Treuner-Lange, K. Aguiluz, C. van der Does, N. Gómez-Santos, A. Harms, D. Schumacher, P. Lenz, M. Hoppert, J. Kahnt, J. Muñoz-Dorado, and L. Søgaard-Andersen, “Pom Z, a Par A-like protein, regulates Z-ring formation and cell division in Myxococcus xanthus ,” Mol Microbiol , vol. 87, pp. 235–253, 2013.
- 6(6) D. Schumacher and L. Søgaard-Andersen, “Regulation of cell polarity in motility and cell division in Myxococcus xanthus ,” Annu Rev Microbiol , vol. 71, pp. 61–78, 2017.
- 7(7) M. A. J. Roberts, G. H. Wadhams, K. A. Hadfield, S. Tickner, and J. P. Armitage, “Par A-like protein uses nonspecific chromosomal DNA binding to partition protein complexes,” Proc Natl Acad Sci U S A , vol. 109, no. 17, pp. 6698–6703, 2012.
- 8(8) D. F. Savage, B. Afonso, A. H. Chen, and P. A. Silver, “Spatially ordered dynamics of the bacterial carbon fixation machinery,” Science , vol. 327, no. 5970, pp. 1258–1261, 2010.