Effect of Local Binding on Stochastic Transport in Ion Channels
I. Kh. Kaufman, W. A. T. Gibby, D. G. Luchinsky, P. V. E. McClintock

TL;DR
This paper explores how local binding effects influence ionic Coulomb blockade in ion channels, revealing geometry-dependent shifts and ion-ion interactions that affect ion permeation and resonance behavior.
Contribution
It introduces a correction for local site binding in Coulomb blockade models, explaining observed ion profile splitting and resonance shifts in ion channels.
Findings
Local binding causes geometry-dependent shifts in conduction points.
Ion-ion repulsion explains Ca$^{2+}$ profile splitting.
The model aligns with Brownian dynamics simulation results.
Abstract
Ionic Coulomb blockade (ICB) is an electrostatic phenomenon recently discovered in low-capacitance ion channels/nanopores. Depending on the fixed charge that is present, ICB strongly and selectively influences the ease with which a given type of ion can permeate the pore. The phenomenon arises from the discreteness of the charge-carriers, the dielectric self-energy, an electrostatic exclusion principle, and sequential pore neutralization, and it manifests itself strongly for divalent ions (e.g.\ Ca). Ionic Coulomb blockade is closely analogous to electronic Coulomb blockade in quantum dots. In addition to the non-local 1D Coulomb interaction considered in the standard Coulomb blockade approach, we now propose a correction to take account of the singular part of the attraction to the binding site (i.e.\ local site binding). We show that this correction leads to a…
Click any figure to enlarge with its caption.
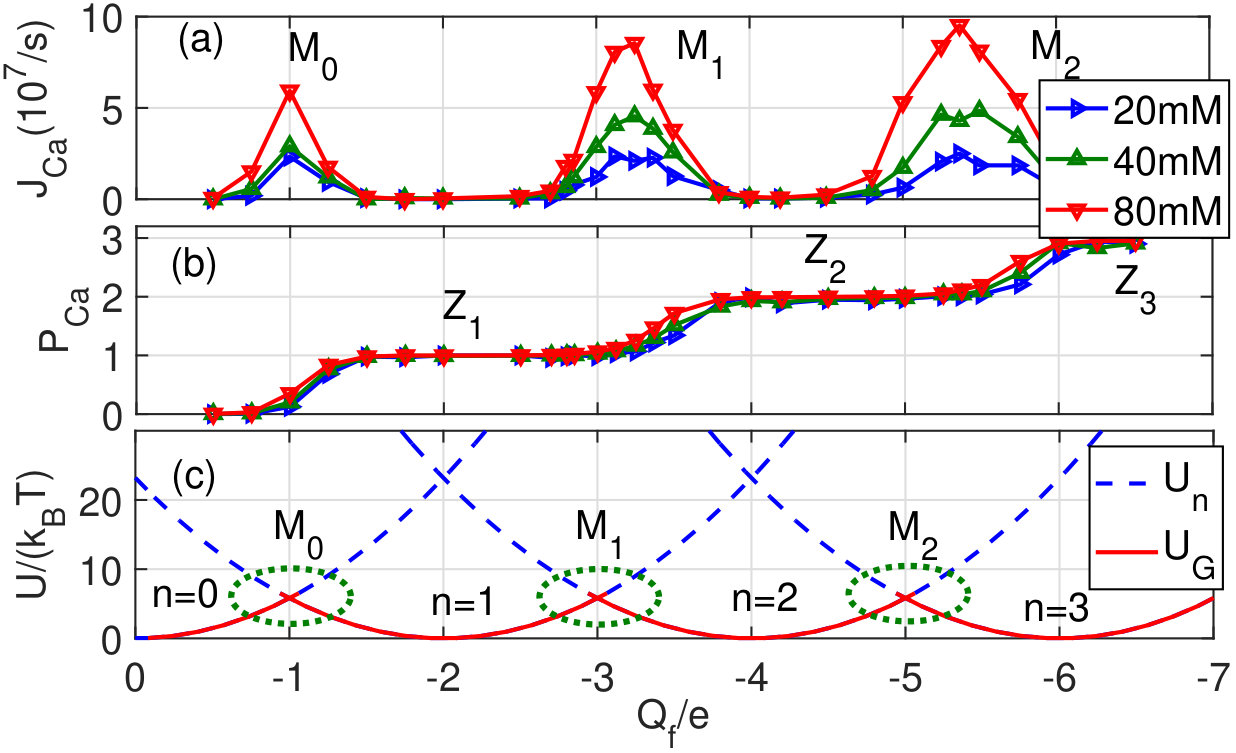 Figure 1
Figure 1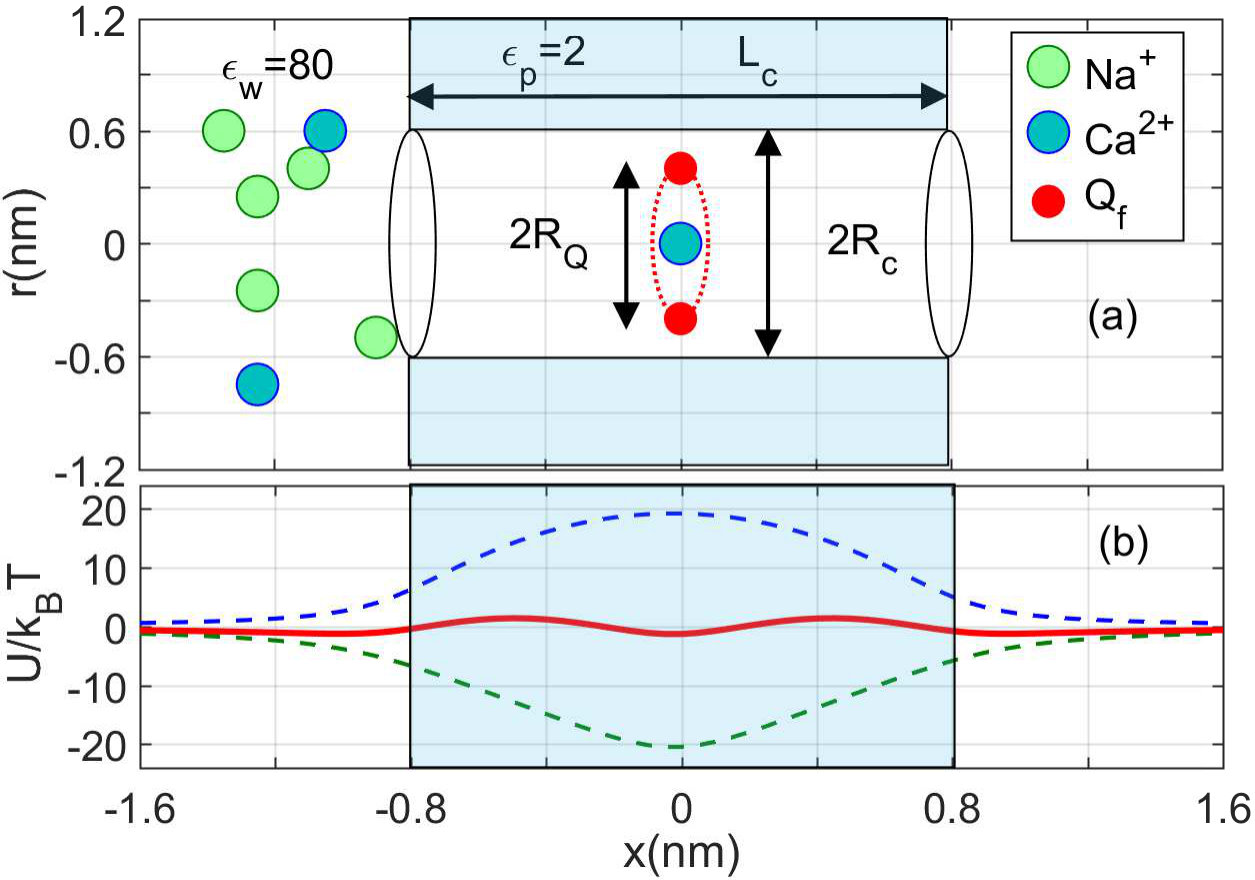 Figure 2
Figure 2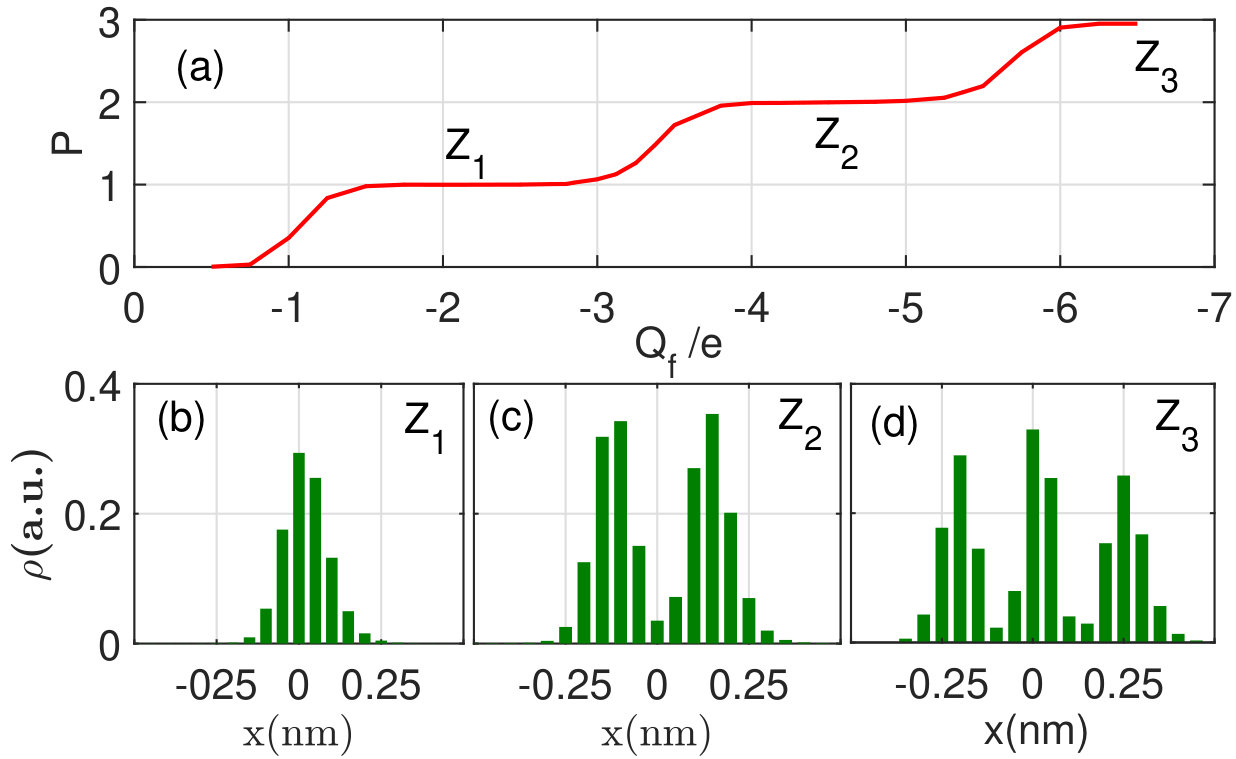 Figure 3
Figure 3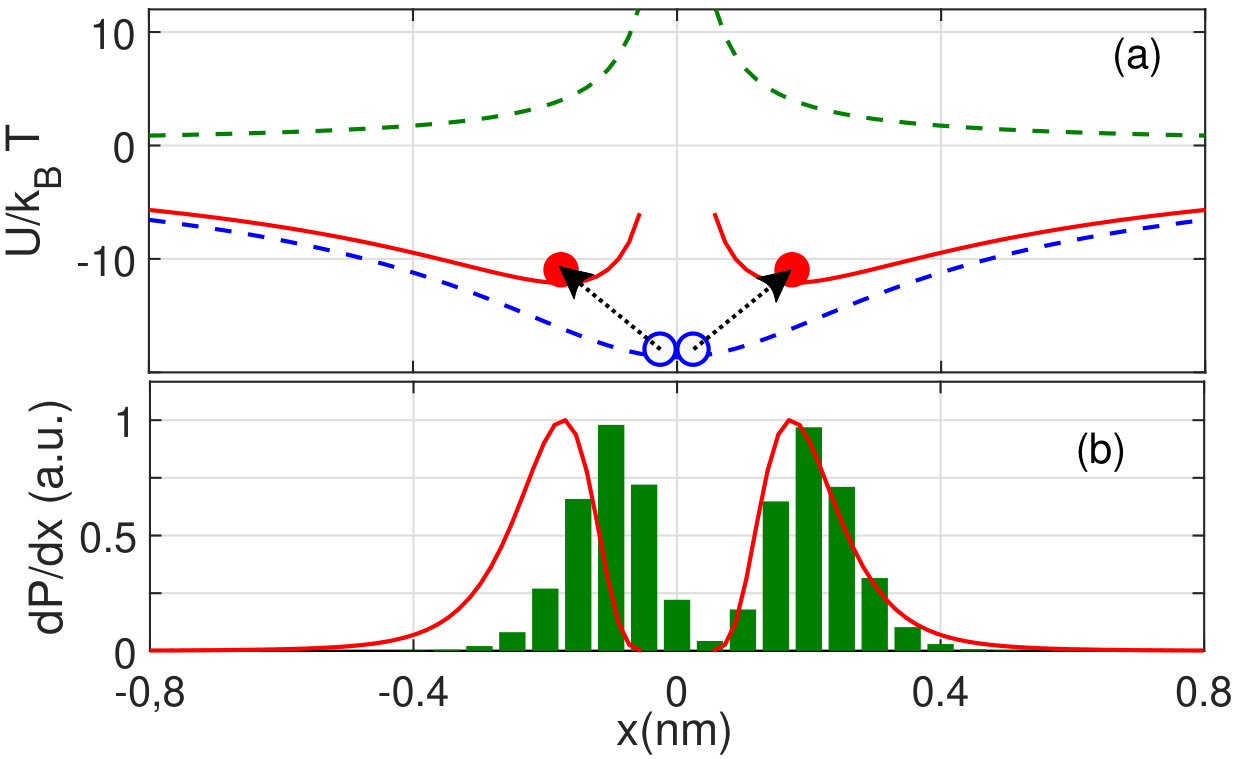 Figure 4
Figure 4Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Effect of Local Binding on Stochastic Transport in Ion Channels
I. Kh. Kaufman1, W. A. T. Gibby1, D. G. Luchinsky12, P. V. E. McClintock1
1Department of Physics, Lancaster University, Lancaster LA1 4YB, UK
Email: [email protected]
2SGT Inc., Greenbelt MD, 20770, USA
Abstract
Ionic Coulomb blockade is an electrostatic phenomenon recently discovered in low-capacitance ion channels/nanopores. Depending on the fixed charge that is present, Coulomb blockade strongly and selectively influences the ease with which a given type of ion can permeate the pore. The phenomenon arises from the discreteness of the charge-carriers and it manifests itself strongly for divalent ions (e.g. Ca2+). Ionic Coulomb blockade is closely analogous to electronic Coulomb blockade in quantum dots. In addition to the non-local 1D Coulomb interaction considered in the standard Coulomb blockade approach, we now propose a correction to take account of the singular part of the attraction to the binding site (i.e. local site binding). We show that this correction leads to a geometry-dependent shift of the single-ion barrierless resonant conduction points M0. We also show that local ion-ion repulsion accounts for a splitting of Ca2+ profiles observed earlier in Brownian dynamics simulations.
I Introduction
Ion channels provide for the selective transport of physiologically important ions (e.g. Na+, K+ and Ca2+) through the bilipid membranes of biological cells. The channels consist of nanopores through proteins embedded in the membrane. Their selectivity for particular ions is determined by the electrostatically-driven stochastic motion of ions within a short, narrow selectivity filter (SF) carrying a binding site with fixed negative charge .
The permeation of ions through the pore is governed by ionic Coulomb blockade (ICB), a phenomenon that manifests itself in low-capacitance systems. It arises as a consequence of the discreteness of the charge-carriers, the dielectric self-energy , an electrostatic exclusion principle, and sequential pore neutralisation as additional ions enter the pore [1, 2, 3]. ICB manifests itself strongly for divalent ions (e.g. Ca2+) [2]. ICB is closely similar to its electronic counterpart in quantum dots [4].
The basic ICB description of the permeation and selectivity of ion channels has already been presented [3]. Here we extend this basic model by the introduction of corrections to allow for the singular part of the attraction of ions to the binding site (i.e. local site binding), in addition to the non-local 1D Coulomb interaction considered in the ICB model [2]. The geometry-dependent shift of the ICB calcium resonant point M0 resulting from this correction leads to a changed threshold for divalent blockade. We will also show that the presence of local (singular) ion-ion repulsion is what leads to the splitting of the Ca2+ axial occupancy profile seen earlier in Brownian dynamics simulations [5, 6].
We start in Sec. II by description of extended electrostatic model of ion channels. In Sec. III we briefly describe the ICB model of permeation and selectivity of calcium/sodium channel. Sec. IV introduces extension accounting for a local binding, followed in Sec. V by consideration of local ion-ion repulsion and resulted multi-ion splitting of calcium profiles. Finally, in Sec. VI we summarize and draw conclusions.
In what follows, with SI units is the permittivity of free space, is proton charge, is Boltzmann’s constant and is the temperature; (I)CB is (ionic) Coulomb blockade, BD is Brownian dynamics, LB is local binding, LR is local repulsion, SF is the selectivity filter and SE is the self-energy. ††The research was supported by the Engineering and Physical Sciences Research Council (EPSRC) UK (grant No. EP/M015831)
II Extended Electrostatic Model of Ion Channel
Fig. 1(a) shows the extended electrostatic model of the SF of a calcium/sodium ion channel, We represent it as a negatively-charged, axisymmetric, water-filled, cylindrical pore of radius nm and length nm through the protein hub in the cellular membrane The -axis is coincident with the channel axis and in the center of channel.
There is a symmetrically-placed, uniformly-charged, rigid ring of negative charge . Extending the earlier model, the radius of this charged ring could be different from the channel radius , corresponding to the charged residues moving partially into the channel. We take both the water and the protein to be homogeneous continua with dielectric constants and , respectively.
Fig. 1(b) illustrates the phenomenon of resonant barrier-less conduction, which is typical of electrostatic models and which occurs when the energy of ion-site attraction balances the dielectric self-energy barrier [7, 8, 2].
This generic electrostatic channel model is similar to that used previously [9, 5, 10, 11]. Details of the model, and its validity and limitations, have already been discussed [6].
III Ionic Coulomb blockade
We consider the stochastic transport of a fully-hydrated Ca2+ ion having charge .
Fig. (2)(a) illustrates the multi-ion Ca2+ conduction bands and (b) shows the corresponding Coulomb staircase in occupancy, as revealed by Brownian dynamics simulations. Steps of this staircase are described by Fermi-Dirac function [2, 3]. The ground state energy diagram for such conductance is plotted in (c). The ICB model [3] states that resonant (barrier-less) conduction points Mn occur when the difference in free energy between states (with ions near the center of the SF) and (channel with ions in the SF plus 1 ion in the bulk) is zero:
[TABLE]
where refers to state , is the potential energy and is the entropy difference. When , there are stable ICB points Zn.
The standard ICB model [2] assumes that with {n} similar ions near the centre is equal to the dielectric self-energy of the excess charge of the SF :
[TABLE]
where is the SF self-capacity.
For simplicity, we consider the first resonant point M0 [6] corresponding to the movement of a single ion through an otherwise empty SF (the transition) so we temporarily ignore the ion-ion interaction term (see Sec. V). Expanding the quadratic form in (2) gives us the following decomposition for ion-related part of potential energy (for )
[TABLE]
where is the ion self-energy , and is the 1D Coulomb ion-site attraction energy [10, 12].
The base position (without the entropy term) for resonant conduction is defined by the condition for barrier-less motion [2, 3]:
[TABLE]
whereas Z corresponds to the SF neutralisation condition. Inclusion of the entropy term leads to a concentration-related shift of the resonance point [2]:
[TABLE]
where Pb is the equivalent bulk occupancy related to the SF volume , is the number density of selected ions in the bulk, and is the reference density; note that for (i.e. for ). For a typical SF geometry condition () corresponds to the concentration [Ca]200mM/l. The dependence of the resonance point’s position on coincides with both simulations and with earlier analytic results [13, 14].
IV Local binding
Next we introduce a local binding (LB) correction by adding the ion-site () 3D -screened Coulomb interaction (see [15]) with energy to the total ion potential energy . It leads to a geometry-dependent shift in the resonance point M:
[TABLE]
where is the dimensionless“SF shape ratio”. For an embedded charge ring (), reduces to . For typical geometries (0.3nm , nm) the correction for LB can be about 0.2.
In summary,
[TABLE]
The “shift-equation” (9) allows us to describe the whole range of ICB phenomenona, as embodied in different shifts of M0 and their possible interference such as a divalent blockade and its dependence on [16, 2], or concentration-related shifts of the Coulomb staircase –
- •
The standard ICB optimal conduction point M defines the barrier-less point for ion of charge with at the standard bulk density ().
- •
The LB shift accounts for the real shape of the SF and/or for .
- •
The concentration-related shift describes the influence of .
- •
The interference between and could explain observable differences in divalent blockade thresholds between equally-charged (D/E) mutants of the calcium [16] or bacterial sodium channels [18].
The axial potential energy profiles for the ICB model can be derived from the 1D Coulomb gas approximation [10, 12]:
[TABLE]
where is the dielectric self-energy profile and is the ion-site binding energy. Profile for LB correction can be calculated from Coulomb’s law [15]:
[TABLE]
V Local ion-ion repulsion
In the above analytics we ignored ion-ion repulsion, but here we take it explicitly into account and work out its consequences. For simplicity we consider 2 similar ions located symmetrically () around at . In such a case, the 1D Coulomb ion-ion repulsion and the additional local 3D Coulomb repulsion are respectively:
[TABLE]
The total energy of an ion inside the SF, allowing for both LB and LR, can be expressed as:
[TABLE]
Hence for the neutralization point Zn , the energy profile is defined by local components only.
Fig. 3(a) shows that the energy function calculated according to (13) has two symmetrical off-center minima defining a splitting of the Ca2+ occupancy profile. Thus, by taking account of LB and of the local ion-ion repulsion in the SF, we arrive at an explicit, self-consistent, analytic explanation of the splitting of the multi-ion occupancy profiles observed earlier in Brownian dynamics simulations [5, 6, 17], in self-consistent numerical solutions of the Poisson equation [17], and in analytic non-self-consistent calculations [19, 20]. This splitting leads to significant increases in ionic energy (as indicated by arrows) and eventually to knock-on escape. Note that the “binding points” () are unconnected with any physical binding sites different from the main -related site. This situation can be described as “’virtual sites’ or self-organisation of ions inside the SF (see also [11, 21]) .
Fig. 3(b) compares the Ca2+ occupancy profile returned by BD simulations (green histogram) with that estimated from the potential energy profile as (red solid line). The satisfactory agreement obtained can be regarded as confirming the consistency of our model.
Fig. 4 shows the evolution of the Ca2+ profile with varying as found from BD simulations [17, 22].
Fig. 4(a) shows the simulated calcium occupancy of the SF vs for bath concentration [Ca]=80mM (see Fig.1(b)), demonstrating the standard Coulomb staircase shape with Fermi-Dirac steps. The single-ion Z, double-ion Z and triple-ion Z blockade points are indicated.
Fig. 4 (b)-(d) represent BD-simulated profiles for different values. In (b) there is an (unsplit) single-ion occupancy profile for the Z1 point, which appears due to ICB and LB of the ion to .
The local repulsion provides for self-organisation of the ions and splitting of the occupancy profile for the double-ion Z2 point (c) and triple-ion Z3 point (d). Note that, for intermediate values of (e.g. between Z1 and Z2), the {n}-state will be mixed (time shared), providing averaged profiles having an arbitrary number of peaks.
VI Conclusions
The effect of local binding on conduction and occupancy (analytic result (9)) is found to agree with Brownian dynamics simulations, thus putatively accounting for how the position of the resonant point M0 is influenced by the radius of the Glutamate/Aspartate ring in NaChBac channels and their mutants [23, 18, 24].
The local binding and local repulsion lead to corrections of about 20 and to an observable splitting of the Ca2+ occupancy profiles. The splitting of the Ca2+ profile is found in reasonable quantitative agreement with the results of BD simulations [6].
These results are also applicable to artificial sub-nm pores [25].
Acknowledgment
We are grateful to M. Di Ventra, R. S. Eisenberg, O. A. Fedorenko, C. Guardiani, S. K. Roberts, and A. Stefanovska for comments and useful discussions.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1[1] M. Krems and M. Di Ventra, “Ionic Coulomb blockade in nanopores,” J. Phys. Condens. Matter , vol. 25, p. 065101, 2013.
- 2[2] I. K. Kaufman, P. V. E. Mc Clintock, and R. S. Eisenberg, “Coulomb blockade model of permeation and selectivity in biological ion channels,” New J. Phys. , vol. 17, no. 8, p. 083021, 2015.
- 3[3] I. Kaufman, W. Gibby, D. Luchinsky, P. Mc Clintock, and R. Eisenberg, “Coulomb blockade oscillations in biological ion channels,” in Proc. 23rd Intern. Conf. on Noise and Fluctuations (ICNF), Xian . IEEE Conf. Proc., June 2015, p. doi: 10.1109/ICNF.2015.7288558.
- 4[4] C. W. J. Beenakker, “Theory of Coulomb-blockade oscillations in the conductance of a quantum dot,” Phys. Rev. B , vol. 44, no. 4, pp. 1646–1656, 1991.
- 5[5] B. Corry, T. W. Allen, S. Kuyucak, and S. H. Chung, “Mechanisms of permeation and selectivity in calcium channels,” Biophys. J. , vol. 80, no. 1, pp. 195–214, 2001.
- 6[6] I. K. Kaufman, D. G. Luchinsky, R. Tindjong, P. V. E. Mc Clintock, and R. S. Eisenberg, “Multi-ion conduction bands in a simple model of calcium ion channels,” Phys. Biol. , vol. 10, no. 2, p. 026007, 2013.
- 7[7] G. Eisenman and R. Horn, “Ionic selectivity revisited: the role of kinetic and equilibrium processes in ion permeation through channels,” J. Membrane Biol. , vol. 76, no. 3, pp. 197–225, 1983.
- 8[8] S. O. Yesylevskyy and V. N. Kharkyanen, “Barrier-less knock-on conduction in ion channels: peculiarity or general mechanism?” Chem. Phys. , vol. 312, pp. 127–133, 2005.