The joint influence of competition and mutualism on the biodiversity of mutualistic ecosystems
Carlos Gracia-L\'azaro, Laura Hern\'andez, Javier Borge-Holthoefer,, and Yamir Moreno

TL;DR
This paper introduces a multilayer network framework to analyze how competition and mutualism jointly influence biodiversity in ecosystems, revealing complex interactions that affect species coexistence and ecosystem stability.
Contribution
It develops a novel multilayer network approach that explicitly models both mutualistic and competitive interactions, moving beyond mean-field approximations in ecological network analysis.
Findings
Mutualism's effect varies between specialist and generalist species.
Biodiversity exhibits a non-trivial profile depending on competition and mutualism levels.
Considering both interactions simultaneously alters understanding of ecosystem stability.
Abstract
Relations among species in ecosystems can be represented by complex networks where both negative (competition) and positive (mutualism) interactions are concurrently present. Recently, it has been shown that many ecosystems can be cast into mutualistic networks, and that nestedness reduces effective inter-species competition, thus facilitating mutually beneficial interactions and increasing the number of coexisting species or the biodiversity. However, current approaches neglect the structure of inter-species competition by adopting a mean-field perspective that does not deal with competitive interactions properly. Here, we introduce a framework based on the concept of multilayer networks, which naturally accounts for both mutualism and competition. Hence, we abandon the mean field hypothesis and show, through a dynamical population model and numerical simulations, that there is an…
Click any figure to enlarge with its caption.
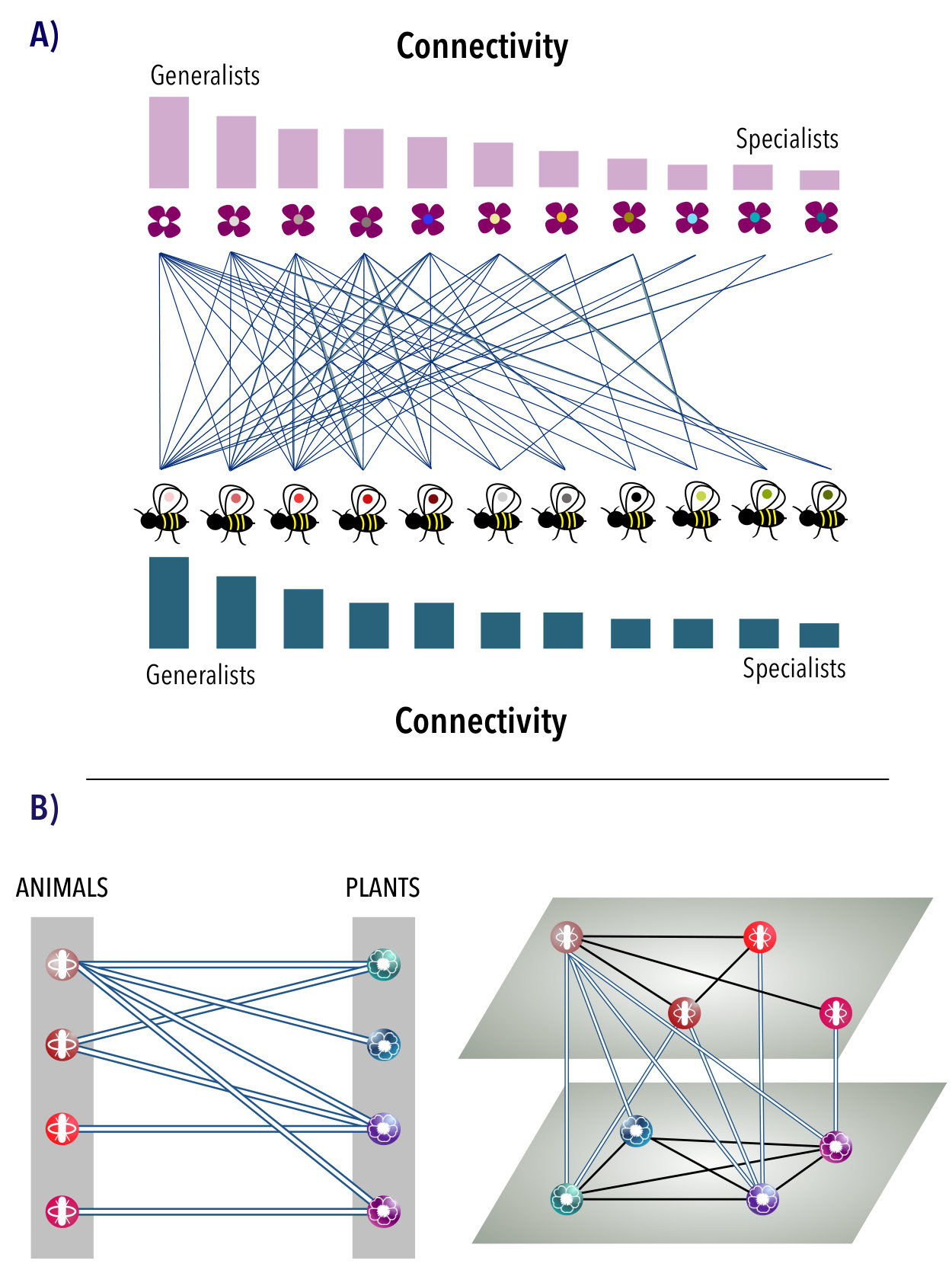 Figure 1
Figure 1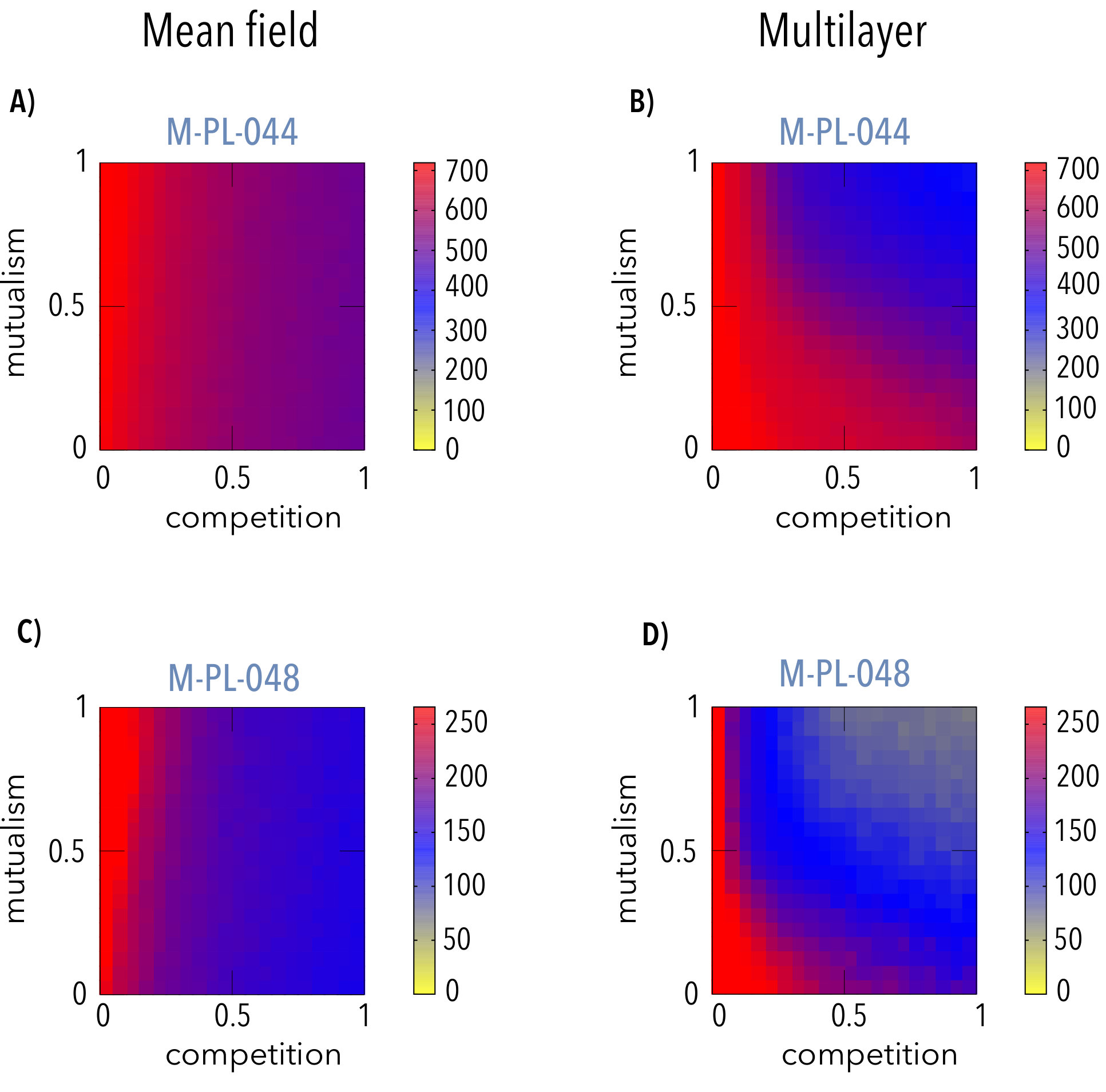 Figure 2
Figure 2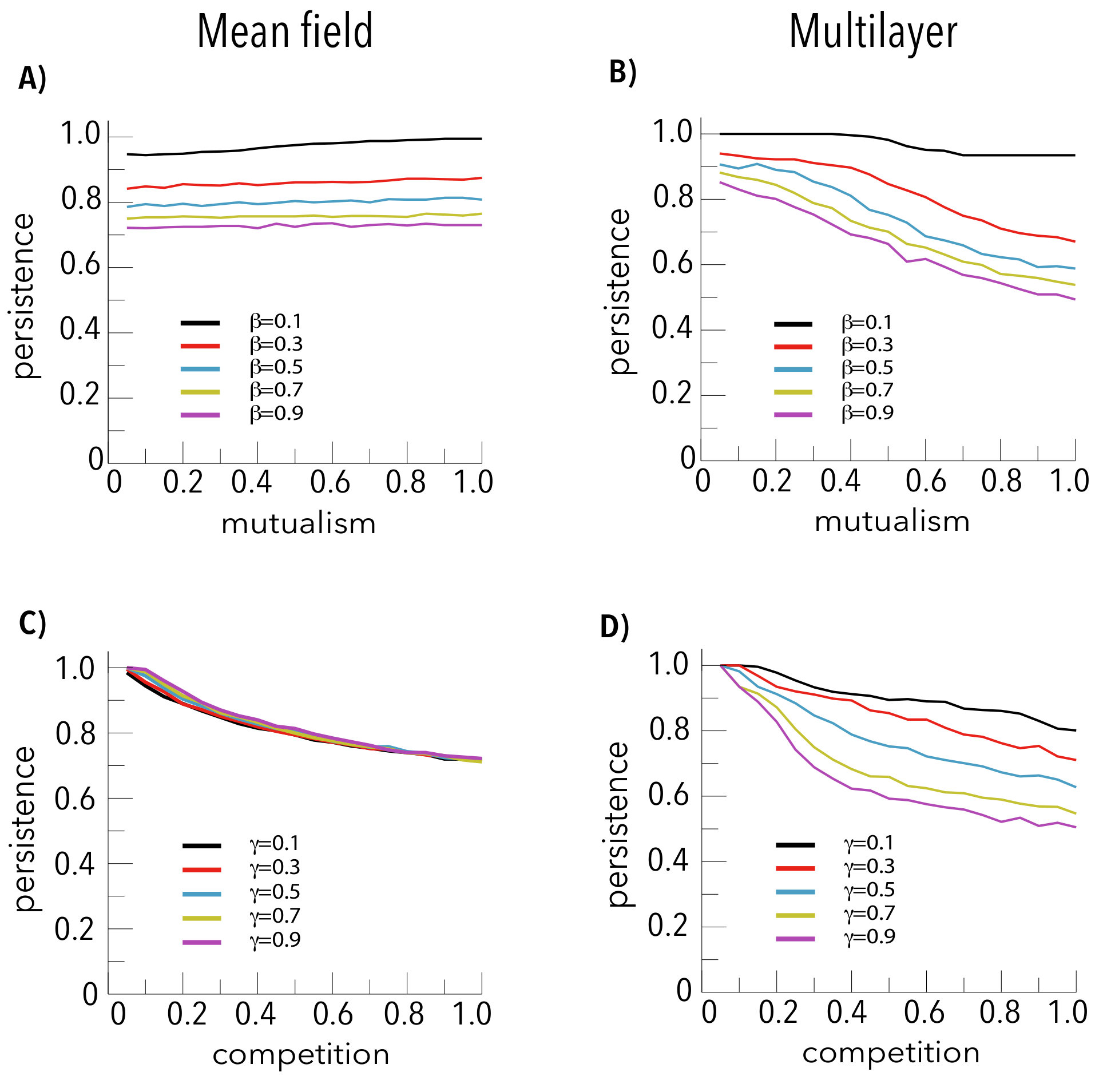 Figure 3
Figure 3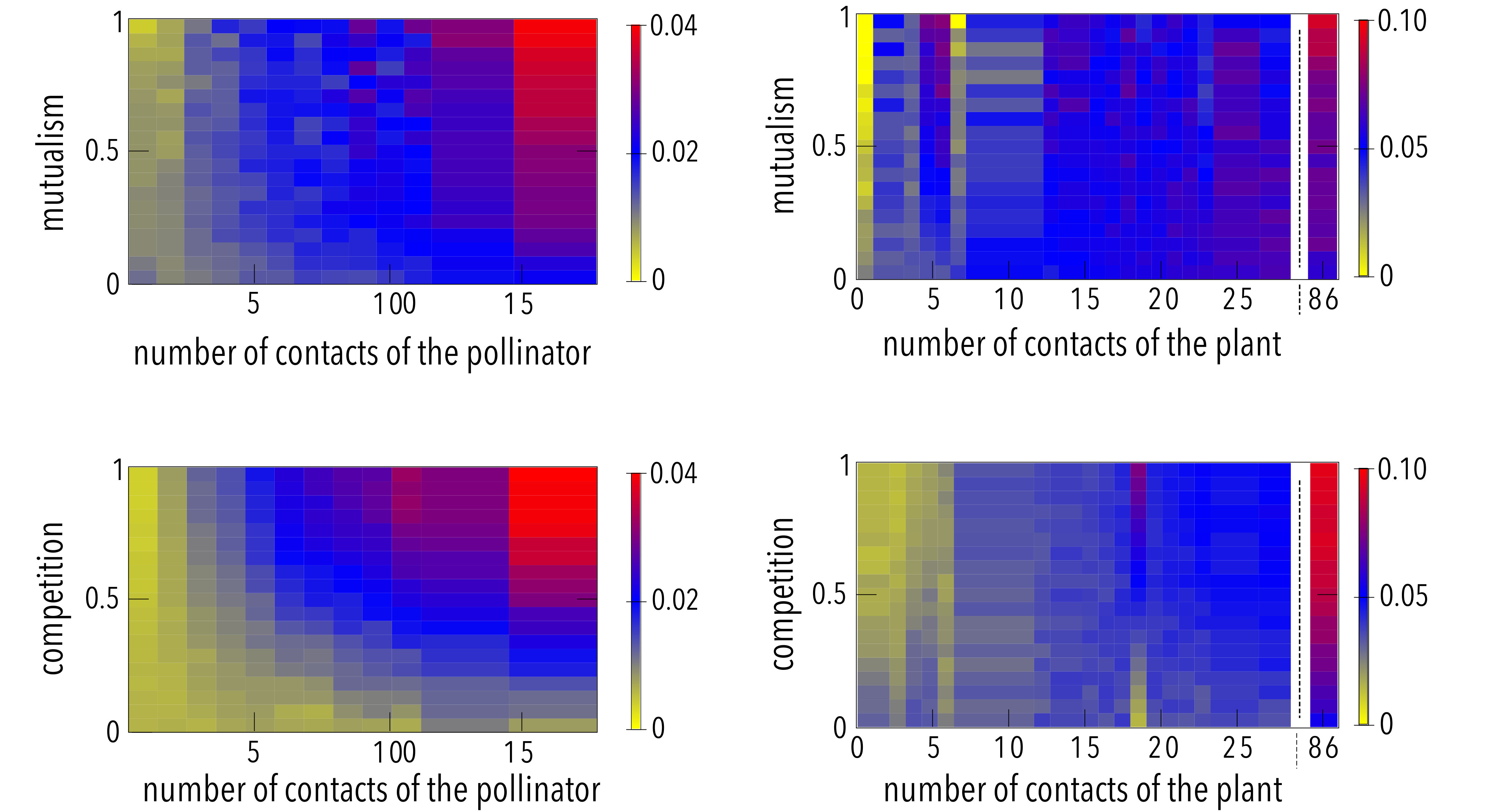 Figure 4
Figure 4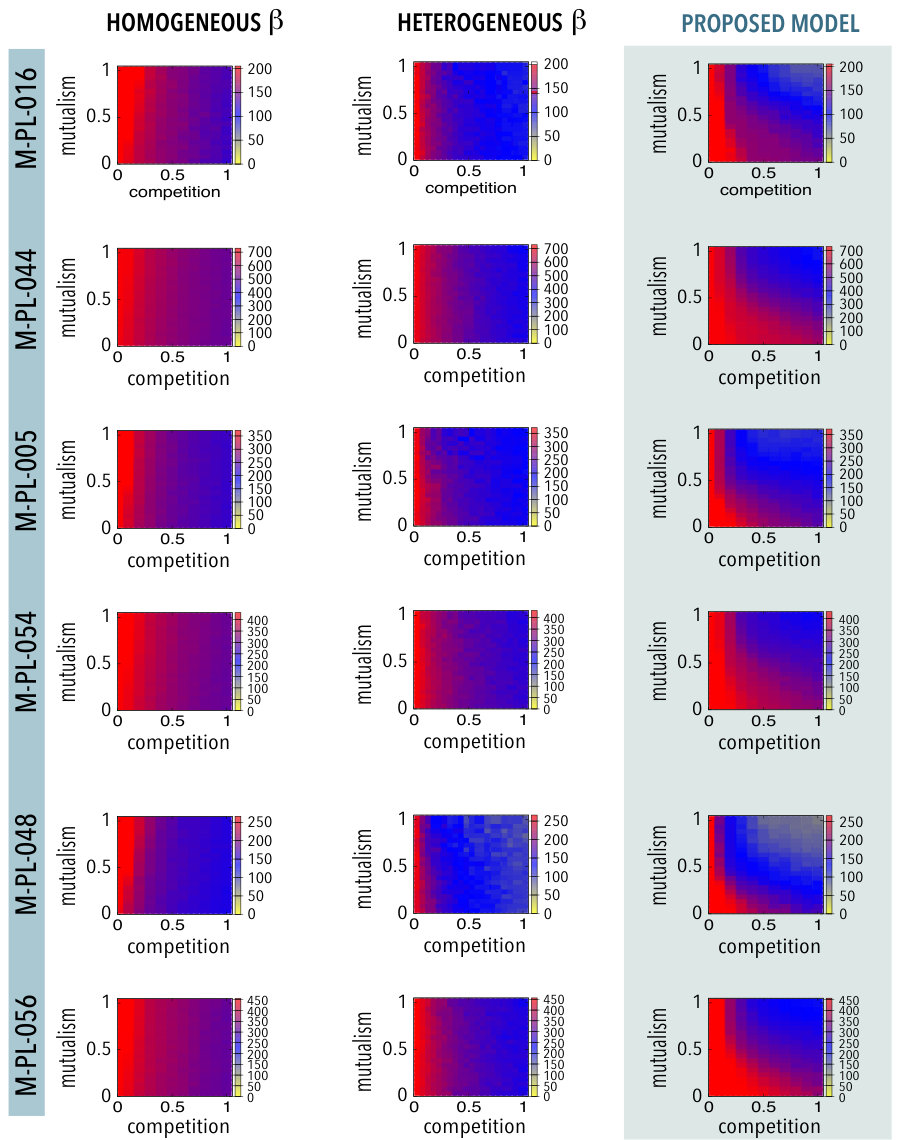 Figure 5
Figure 5| network | species | plants | pollinators | location | reference |
|---|---|---|---|---|---|
| M-PL-016 | 205 | 26 | 179 | Doñana Nat. Park, Spain | [21] |
| M-PL-044 | 719 | 110 | 609 | Amami-Ohsima Island, Japan | [22] |
| M-PL-005 | 371 | 96 | 205 | Pikes Peak, Colorado, USA | [23] |
| M-PL-054 | 431 | 113 | 318 | Kyoto, Japan | [24] |
| M-PL-048 | 266 | 30 | 236 | Denmark | [25] |
| M-PL-056 | 456 | 91 | 365 | Mt. Kushigata, Japan | [26] |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
\JournalInfo\Archive\PaperTitle
The joint influence of competition and mutualism on the biodiversity of mutualistic ecosystems \AuthorsCarlos Gracia-Lázaro1, Laura Hernández2, Javier Borge-Holthoefer3, and Yamir Moreno1,4,5*
\KeywordsMutualistic Ecosystems — Multilayer Networks — Biodiversity – Nestedness \AbstractRelations among species in ecosystems can be represented by complex networks where both negative (competition) and positive (mutualism) interactions are concurrently present. Recently, it has been shown that many ecosystems can be cast into mutualistic networks, and that nestedness reduces effective inter-species competition, thus facilitating mutually beneficial interactions and increasing the number of coexisting species or the biodiversity. However, current approaches neglect the structure of inter-species competition by adopting a mean-field perspective that does not deal with competitive interactions properly. Here, we introduce a framework based on the concept of multilayer networks, which naturally accounts for both mutualism and competition. Hence, we abandon the mean field hypothesis and show, through a dynamical population model and numerical simulations, that there is an intricate relation between competition and mutualism. Specifically, we show that when all interactions are taken into account, mutualism does not have the same consequences on the evolution of specialist and generalist species. This leads to a non-trivial profile of biodiversity in the parameter space of competition and mutualism. Our findings emphasize how the simultaneous consideration of positive and negative interactions can contribute to our understanding of the delicate trade-offs between topology and biodiversity in ecosystems and call for a reconsideration of previous findings in theoretical ecology, as they may affect the structural and dynamical stability of mutualistic systems.
Ecosystems research, and particularly the study of mutualistic communities (like plant and pollinators or plant and seed dispersers), has recently witnessed a major conceptual leap. This has been possible mainly due to two factors: the implementation of more complex dynamical population models that have allowed to go beyond the linear random matrix interaction model of May[1]; and the introduction of a networked perspective that uncovered the consequences of certain structural arrangements at the system level[2, 3, 4, 5], with nestedness at the forefront [6, 7, 8, 9]. Nestedness is a widespread property of mutualistic ecosystems. It allows to quantify key interactions in these systems, particularly the so-called mutualistic interactions, where specialist and generalist species of two guilds (having a small and a large number of inter-guild interactions, respectively, see Figure 1a) facilitate their mutual coexistence. The structure and the dynamics (as given by the persistence or biodiversity of species) of these ecosystems have been shown to be intimately connected[5].
Additionally, nestedness has emerged as a rather common mesoscale pattern in complex bipartite networks, suggesting that other systems, beyond ecological communities[10, 11, 12, 13, 14, 15], might follow a similar competition-minimization principle[5]. However, both a bipartite representation and the nestedness of the system fail to incorporate intra-guild (competitive) links –so far accounted for within a mean-field description–, precluding current research to develop an even more realistic framework. Although some alternative representations have been recently suggested[16, 17, 18], they still fall short when it comes to inspect the feedback between structure and dynamics of mutualistic systems. In parallel, recent advances in data collection suggest that considering actual (rather than probabilistic) information on intra-guild connectivity will be feasible soon[19]. It is then imperative to properly deal with both positive (mutualistic) and negative (competitive) interactions in a way that naturally allows to plug in dynamical population models. To this end, here we capitalize on the recently developed framework of multilayer networks[20], and show that it is possible to encode within a unique topological representation, both kinds of interactions. In turn, this allows us to consider, through analytical and numerical results, how the biodiversity of the system varies as a function of the intensities of mutualism and competition in the system.
In order to build the multilayer representation of a mutualistic ecosystem, we exploit the implicit information contained in the bipartite matrix: the projections onto the subspace of species A (animals) and species P (plants) yield two hidden weighted networks that reveal the existent intra-guild interactions, see Figure 1B, which are encoded in the adjacency matrices associated to each layer. These topological relations can be used to model inter-species competition beyond the mean-field approach, as they explicitly take into account the actual architecture of intra-layer interactions. Note that in this representation, mutualistic interactions are accounted for by the inter-layer connections between elements of A and P, see Figure 1b. We next investigate the influence of the network structure on the persistence of species of the mutualistic ecosystem. The main point of interest is whether the competitive interactions, as given by the multilayer topology, actually convey substantive dynamical changes. To this end, we implement a population dynamical model that capitalizes on the one introduced by Bastolla et al. [5], but constrained by the multilayer architecture. We study the variation of biodiversity (e.g., the number of species at the steady state of the population dynamics) on a diverse set of real ecological communities[21, 22, 23, 24, 25, 26] (see also the Supplementary Information), for a wide range of parameters of the model.
Let’s assume that the mutualistic community consists of species of plants and species of animals (pollinators or seed-dispersers); the biodiversity is denoted by . We denote by the abundance of the plant species , and by its intrinsic growth rate. Similarly, animals’ parameters and variables are represented by the superscript . The mutualistic relationships (i.e., the inter-layer connections) are given by a bipartite matrix, , with if animal species pollinates the plant species , and otherwise. The biomass of the pollinators of a given plant species is thus . On the other hand, the intra-layer relationships represent the resources that are shared by species of the same guild. Therefore, the biomass of the pollinators shared by two plant species is . Finally, the relative abundance of a given plant evolves according to:
[TABLE]
The first term of this equation represents the intrinsic growth of species without considering saturation and the second term refers to the intra-specific competition term (saturation), which can be interpreted in terms of a carrying capacity in the absence of competing species. The third term of Eq. (1) accounts for the intra-guild inter-specific competition. Here, the competition between two plant species () is weighted according to the incidence of the biomass of shared pollinators, , in the biomass of pollinators of each plant species, . Lastly, the fourth term in Eq. (1) gives the contribution of mutualism to the abundance of plant species , being the Holling term that imposes a limit to the mutualistic effect. The intensities of competition and mutualism and respectively, constitute the parameter space that we investigate. The corresponding equation for the abundance of pollinators is equivalent to Eq. (1) but interchanging superscripts by and vice versa (see Supplementary Information).
We numerically solved the system of equations describing the abundances of plants and animals (see Methods). Figure 2 compares results obtained for the system’s biodiversity when the single (panels a and c) and multilayer (panels b and d) population models are implemented in two real networks (M_PL_044[22] in panels a and b; and M_PL_048[25] in panels c and d) as a function of the intensities of mutualism and competition. The single layer population model corresponds to the mean-field version of the competition term[5] while the mutualistic term does contain the information about the structure of the real network. On the contrary, the multilayer setup naturally deals with the structure of the competitive interactions. As seen in the figure (see also the Supplementary Information for results corresponding to other real networks), the biodiversity of the system is greatly affected by intra-guild competition. In the mean-field approach, where all species compete on equal grounds with all the others of the same guild, the persistence of biodiversity in the real systems does not depend on . Conversely, when the structure of the real network is introduced in the inter-species competition term, the region of structural stability depends, non-trivially, on both parameters, and .
The results shown in Figure 2 may appear counterintuitive. Indeed, if mutualistic interactions were to reduce effective competition and increase biodiversity[5], one should expect that the boundary separating the region where all species survive (coded in red in Fig. 2) from the one where biodiversity diminishes (coded in blue in Fig. 2) would behave as a monotonous growing curve, which is clearly not the case. Figure 3 illustrates this point. As expected, the biodiversity is a decreasing function of the competition parameter () in both settings when the mutualism intensity () is fixed (panels c and d). Note, additionally, that in the mean-field case, the persistence is independent of , as also shown in Figure 2.Remarkably enough, when the competition term takes into account the actual interactions in the multilayer frame, the same decreasing behavior for the persistence of species is also observed when the intensity of competition is kept constant and the extent of mutualism is varied, see panel b in Fig. 3, except for very low values, where competition may be neglected and the biodiversity does not vary with mutualism. This paradoxical result is due to the joint action of the mutualistic and the inter-species competition terms, which lead to differences on the way the abundance of generalist and specialist species evolves (see Methods for an heuristic argument and the Supplementary Information). The latter selective influence of both mutualism and competition on specialists’ and generalists’ species is illustrated in Figure 4, where we represent the relative abundance of the species as a function of the species’ connectivity and the intensities of mutualism (top panels, in which the competition is fixed) and competition (bottom panels, in which the mutualism is fixed). The results are clear-cut: it turns out that species with higher degrees remain relatively more abundant than those with lower degrees when there is an increase of the strength of either mutualistic or competitive interactions.
The results that come out from the approach here adopted unveil the important role played by the network structure of the inter-species competition term on the biodiversity of the system. When this interaction is treated in a mean-field fashion, the biodiversity persists for any intensity of mutualism, as long as the interspecific competition remains under a certain value the frontiers indicating the loss of biodiversity are vertical in panels a and c of Fig. 2. On the contrary, when considering the network structure in the mutualistic and in the intra-guild competition term, the region of the parameters space where the biodiversity persists depends non trivially on the intensity of both. In other words, increasing the intensity of mutualism (for a given network) does not necessarily increase biodiversity. Indeed, depending on the intensity of competitive interactions, higher levels of mutualism are detrimental for the survival of the (specialist) species. Therefore, our findings lessen the relevance of the widespread belief that mutualism diminishes competition, thus enhancing biodiversity[5] when it comes to analyze the persistence of species in mutualistic systems. The latter is only roughly valid for weak competition levels, when increasing mutualism is not detrimental for the biodiversity (see Figure 3).
However, our results do not imply that mutualism is not relevant in order to explain the existence of large complex ecosystems. Instead, the careful treatment of the structure of the interactions provides a better understanding of the subtle trade-offs between competition and mutualism. In Ref.[28] an indirect mechanism for cooperation via the interaction with a common counterpart was discussed. Here we show, in addition, that the asymmetry of the competition term between a generalist and a specialist, induced by mutualism, favors the generalist species. This explains why when the intensity of the mutualistic interactions increases, biodiversity may diminish through an important loss of specialists species in favor of the increase of the population of the generalist ones. Moreover, we have checked that our conclusions are indeed due to the structure of the inter-species competition term and not to the fact that the intensity of this competition is not homogeneous. That is, one might think that similar results would be obtained in a mean-field treatment but using heterogeneous intensities for the inter-species competition term. To show that the latter is not the case, we simulated a system in which the inter-species interaction is such that each species interacts with all the others but with a non homogeneous intensity drawn from a distribution. The results (details can be found in the Supplementary Information) indicate that this heterogeneity is not enough to reproduce the same patterns of biodiversity obtained when the projected, multilayer network representation is considered.
In summary, we have introduced a multilayer approach to the study of mutualistic ecosystems that allows to properly consider competitive interactions among species. Numerical simulations of a dynamical population model that is coupled to the data-driven multilayer architecture revealed that the role of mutualistic interactions in maintaining the system’s biodiversity is not as important and as differentiating as previously thought[5, 27]. Instead, we have reported that both mutualism and competition play a complementary role. Strikingly, we have shown that, contrary to what one would have expect, when the level of competition is high, the biodiversity of the system is higher for lower mutualistic intensities and indeed, increasing mutualism maybe detrimental for the species persistence. In light of the present results, there are a number of further questions that remain to be explored, including whether there is an optimal value of mutualism (and competition) at which the system maximizes biodiversity and its dependency with the nestedness of the system. Considering the multilayer approach introduced here would helpfully provide more realistic grounds to tackle these and related challenges.
Methods
Equation for species A.
The dynamical equation that describes the evolution of the abundance of animal species is equivalent to Eq. (1) in the main text, i.e.,
[TABLE]
Numerical Simulations of the Model.
We numerically solve the system of equations (1) and (2), using the matrices that correspond to different real systems (see also Supplementary Information), namely, M_PL_016[21], M_PL_044[22], M_PL_005[23], M_PL_054[24], M_PL_048[25] and M_PL_056[26]. Each simulation starts from random initial conditions of the relative abundances. Following Ref.[5, 27] we take the values of from a uniform distribution in the interval [0.9, 1.1], the intra-species competition is fixed to and the Holling term is . With these parameters, we study the system varying the value of the intensity of the inter-species competition and mutualistic terms, and , respectively. For simplicity, we assumed that all the intervening parameters take the same values for plants and animals. Finally, the system is considered to have achieved equilibrium when all the species’ frequencies remain constant. A species is considered to have gone extinct when its relative abundance is lower than .
Evolution of the abundance of generalist and specialist species.
In order to understand how the population dynamics of generalists and specialists is affected by mutualistic and competitive terms, let us first consider the effect of the mutualist term (the fourth term in Eqs. (1) and (2)). This term reads: , where . As is larger for generalists than for specialists, the increasing rate of the former is stronger than that of the latter. In other words, this term favors the increase of generalists species with respect to specialists ones. The analysis of the inter-species competition term (the third term of Eqs. (1) and (2)) is less straightforward. Let us compare the behavior of this term for a generalist and a specialist plant species, of relative abundances and , respectively, in the particular case where the specialist interacts with only one animal species, , of abundance . This means that and . In this extreme case, it is very easy to see that the competing term that enters in Eq. (1) for species reads:
[TABLE]
where the first term corresponds to its competition with the generalist species , and the second term stands for the competition with all the other plant species that share the same pollinator, , of abundance . The corresponding competition term for the generalist is
[TABLE]
where the factor between braces is . The latter expression can be rewritten as:
[TABLE]
The second term of eq. 5, which may include other generalist plants (those that grow faster with ), is reduced by the factors , and in general . Therefore, we can then expect that this unbalance in the corresponding competition terms becomes a supplementary advantage for the generalists, thus reinforcing their growth rate. A similar analysis to estimate the relative importance of the competition term can be done using the properties of projected matrices (see the Supplementary Information).
SUPPLEMENTARY INFORMATION
Multilayer Networks
A graph (i.e., a single-layer network) is a tuple , where stands for the set of nodes and is the set of edges that connect pairs of nodes. The edges of the graph induce a binary relation on that is called the adjacency relation of . This relation can be represented through the adjacency matrix , where indicates the number of links from node to node .
As a particular case, a bipartite graph is a graph whose nodes can be divided into two disjoint sets and (, ) such that every edge connects a node in to one in . The adjacency matrix of a bipartite graph has the form of a block matrix:
[TABLE]
where represents the null matrix, .
A multilayer network is a data structure made of multiple layers, where each layer is a single-layer network. Formally, a multilayer network is a tuple where is a family of graphs (the layers), and () is the set of edges between nodes of different layers.
In order to build a more complete network containing the interactions among species of different guilds as well as intra-guild interactions, we can further exploit the information contained in the bipartite network of mutualistic interactions and consider the projections of the mutualistic interactions onto the guilds of plants () and animals (). This set-up gives us a multilayer network where the guilds constitute the layers, the mutualistic interactions provide the interlayer links and their projections account for the intra-layer links, which in turn represent the number of pollinators (resp., plants) shared by the corresponding plants (resp., pollinators). Accordingly, the resulting multilayer network consists of two layers (), the set of inter-layer links (), and two sets of intra-layer links (, ).
For the sake of clarity, here we adopt the notation commonly used in the biological literature. The mutualistic plant-pollinator interactions (i.e., the inter-layer connections) are given by a bipartite matrix, , with if animal species pollinates the plant species , and otherwise. The projection of this matrix onto the sets of plants is given by:
[TABLE]
where represents the number of pollinators shared by plant species and .
In our dynamical equations, we consider the resources that are shared by species of the same guild. According to Eq. 7, the biomass of the pollinators shared by plant species is given by:
[TABLE]
Equivalently, the respective formulas for pollinators are obtained by interchanging the and superscripts and vice versa, that is,
[TABLE]
[TABLE]
Table 1 shows the main features (number of species, plants and animals), the location of the biotope and the reference of the ecological networks used to numerically solve the differential equations.
Distribution of the competitive term in the mean field approach
Here we explore an alternative formulation of the mean-field treatment. The idea is to explore whether the results obtained are truly due to the new topological constraints (as given by the multilayer network) or if it would be equivalent to introduce the heterogeneity via the intensity of the inter-species competition term. We show below that this is not the case, that is, that the observed trade-off between competition and mutualism when the full network structure is considered in both mutualistic and competitive interactions (Figure 2 of the main text) is not present when a mean-field approximation is adopted for the competition (see also Figure 2 of the main text). Specifically, we have studied the dynamics [5] with a heterogeneous distribution of the competitive strength, that is, the inter-species competition terms are taken from an uniform distribution .
Figure S5 represents the species persistence as a function of the inter-species competition and mutualistic terms (resp., and ). The rows correspond to different real mutualistic networks as indicated. Left panels show the results of integrating the dynamical equations when the competitive term is the same for all the species (given by the horizontal axis). Some of them correspond to those shown in the main text. Additionally, the central panels correspond to the case in which the intensities of the inter-species competition term are taken from a uniform distribution . As it can be seen, a heterogeneous competition implies a lower biodiversity regarding the homogeneous distribution of the competitive term. Finally, the right panels correspond to the model here presented, defined by equations equations (1) and (2) of the main text, and therefore they represent the case in which the competition terms have been weighted proportionally to the shared biomass of pollinators (resp., plants). Again, some of them have been introduced in the main text. A direct comparison of the central and right panels clearly shows that introducing (to some extent artificially) a degree of heterogeneity in the competitive terms cannot reproduce the results obtained when considering the whole network structure characterizing the competitive interactions.
Average relative abundance for animals.
Here we further develop the heuristic argument provided in the Methods section of the main text to explain the effects of the balance between mutualism and inter-specific competition. Let us consider the evolution of the abundance of a plant species , . The factor of eq. (8) involves the abundance of each animal species of the other guild. These abundances are all different because for the animal guild, generalists and specialists also grow faster and slower with , respectively. We will then consider an average relative abundance for animals . Then the competing term, for plants, reads:
[TABLE]
where is the projected matrix on the plant space.
As we can consider two situations:
- •
is a generalist so then, the maximum possible contribution to the competition term is :
- •
is a specialist, so then the maximum contribution to the competition term is :
which shows that the specialists are, on average, more affected by competition than generalists when mutualism increases. The same reasoning holds symmetrically for the animal’s guild.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1[1] May, R.M. Will a Large Complex System be Stable. Nature 238, 413-414 (1972).
- 2[2] Okuyama, T & Holland, J.N. Network structural properties mediate the stability of mutualistic communities. Ecol. Lett. 11, 208-216 (2008).
- 3[3] Thébault E. & Fontaine, C. Stability fo Ecological Communities and the Architecture of Mutualistic and Trophic Networks. Science 329, 853-856 (2010).
- 4[4] Olesen, J.M., Bascompte, J., Dupont, Y.L. & Jordano, P. The modularity of pollination networks. Proc. Natl. Acad. Sci. U.S.A. 104 , 19891-19896 (2007).
- 5[5] Bastolla, U. et al. The architecture of mutualistic networks minimizes competition and increases biodiversity. Nature 458, 1018-1020 (2009).
- 6[6] Jordano, P., Bascompte, J. & Olesen, J.M. Invariant properties in coevolutionary networks of plant–animal interactions. Ecol. lett. 6 , 69-81 (2003).
- 7[7] Bascompte, J., Jordano, P., Melián, C.J. & Olesen, J.M. The nested assembly of plant–animal mutualistic networks. Proc. Natl. Acad. Sci. U.S.A. 100, 9383-9387 (2003).
- 8[8] Bascompte, J., Jordano, P. & Olesen, J.M. Asymmetric coevolutionary networks facilitate biodiversity maintenance. Science 312 431-433 (2006).